

Instability of signaling resolution models of parent-offspring conflict
- PMID: 9539758
- PMCID: PMC22510
- DOI: 10.1073/pnas.95.8.4453
Instability of signaling resolution models of parent-offspring conflict
Abstract
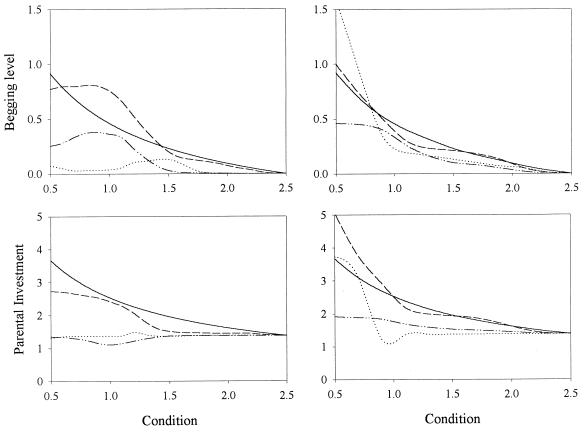
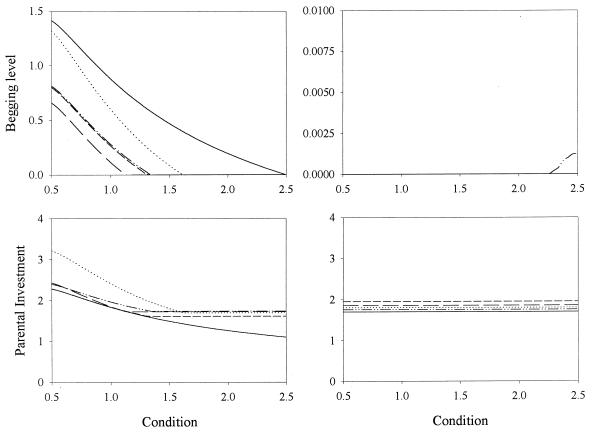
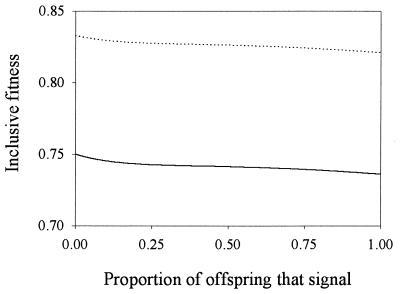
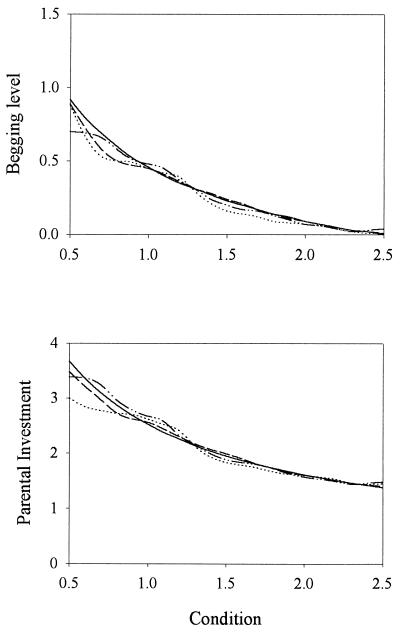
Recent signaling resolution models of parent-offspring conflict have provided an important framework for theoretical and empirical studies of communication and parental care. According to these models, signaling of need is stabilized by its cost. However, our computer simulations of the evolutionary dynamics of chick begging and parental investment show that in Godfray's model the signaling equilibrium is evolutionarily unstable: populations that start at the signaling equilibrium quickly depart from it. Furthermore, the signaling and nonsignaling equilibria are linked by a continuum of equilibria where chicks above a certain condition do not signal and we show that, contrary to intuition, fitness increases monotonically as the proportion of young that signal decreases. This result forces us to reconsider much of the current literature on signaling of need and highlights the need to investigate the evolutionary stability of signaling equilibria based on the handicap principle.
Figures
References
-
- Godfray H C J. Nature (London) 1991;352:328–330.
-
- Godfray H C J. Am Nat. 1991;146:1–24.
-
- Macnair M R, Parker G A. Anim Behav. 1979;27:1202–1209.
-
- Krebs J R, Dawkins R. In: Behavioral Ecology, An Evolutionary Approach. 2nd Ed. Krebs J R, Davies N B, editors. Oxford: Blackwell Scientific; 1984. pp. 380–402.
-
- Harper A B. Am Nat. 1986;128:99–114.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
