

Activating mutations in the extracellular domain of the fibroblast growth factor receptor 2 function by disruption of the disulfide bond in the third immunoglobulin-like domain
- PMID: 9539778
- PMCID: PMC22530
- DOI: 10.1073/pnas.95.8.4567
Activating mutations in the extracellular domain of the fibroblast growth factor receptor 2 function by disruption of the disulfide bond in the third immunoglobulin-like domain
Abstract
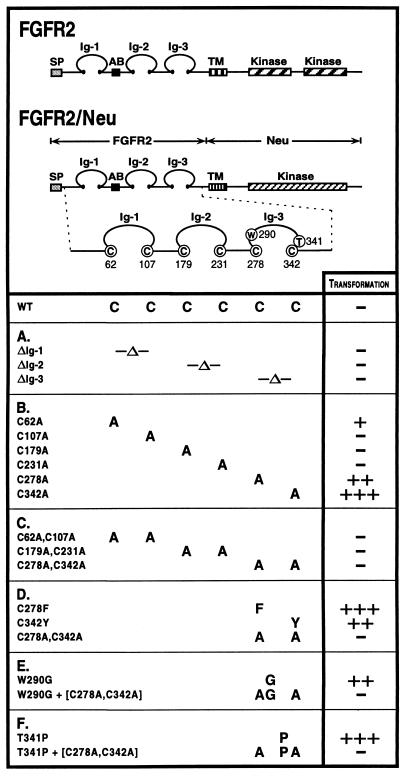
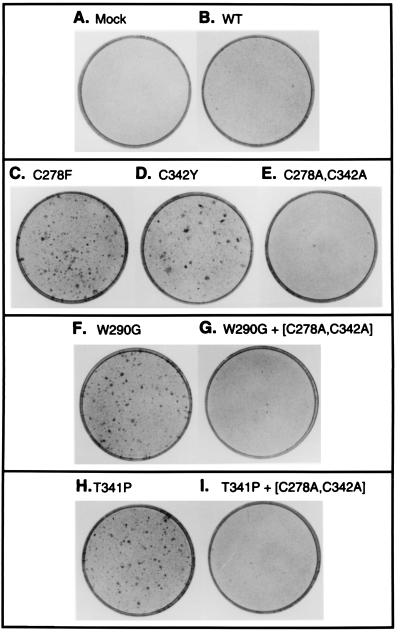
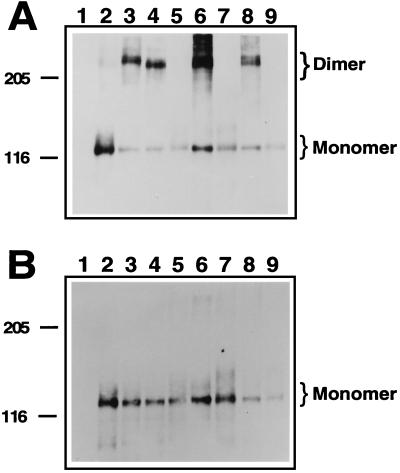
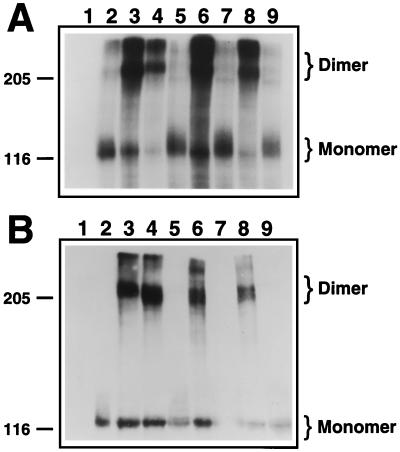
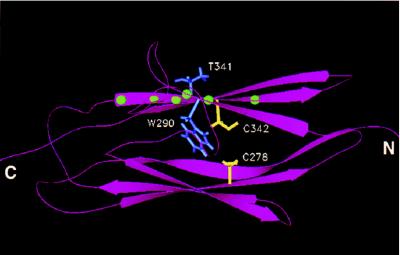
Multiple human skeletal and craniosynostosis disorders, including Crouzon, Pfeiffer, Jackson-Weiss, and Apert syndromes, result from numerous point mutations in the extracellular region of fibroblast growth factor receptor 2 (FGFR2). Many of these mutations create a free cysteine residue that potentially leads to abnormal disulfide bond formation and receptor activation; however, for noncysteine mutations, the mechanism of receptor activation remains unclear. We examined the effect of two of these mutations, W290G and T341P, on receptor dimerization and activation. These mutations resulted in cellular transformation when expressed as FGFR2/Neu chimeric receptors. Additionally, in full-length FGFR2, the mutations induced receptor dimerization and elevated levels of tyrosine kinase activity. Interestingly, transformation by the chimeric receptors, dimerization, and enhanced kinase activity were all abolished if either the W290G or the T341P mutation was expressed in conjunction with mutations that eliminate the disulfide bond in the third immunoglobulin-like domain (Ig-3). These results demonstrate a requirement for the Ig-3 cysteine residues in the activation of FGFR2 by noncysteine mutations. Molecular modeling also reveals that noncysteine mutations may activate FGFR2 by altering the conformation of the Ig-3 domain near the disulfide bond, preventing the formation of an intramolecular bond. This allows the unbonded cysteine residues to participate in intermolecular disulfide bonding, resulting in constitutive activation of the receptor.
Figures
References
-
- Johnson D E, Williams L T. Adv Cancer Res. 1993;60:1–41. - PubMed
-
- Partanen J, Vainikka S, Alitalo K. Philos Trans R Soc London Ser B. 1993;340:297–303. - PubMed
-
- Ullrich A, Schlessinger J. Cell. 1990;61:203–212. - PubMed
-
- Spivak-Kroizman T, Lemmon M A, Dikic I, Ladbury J E, Pinchasi D, Huang J, Jaye M, Crumley G, Schlessinger J, Lax I. Cell. 1994;79:1015–1024. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
