

Dehydroepiandrosterone: a potential signalling molecule for neocortical organization during development
- PMID: 9539798
- PMCID: PMC22550
- DOI: 10.1073/pnas.95.8.4678
Dehydroepiandrosterone: a potential signalling molecule for neocortical organization during development
Abstract
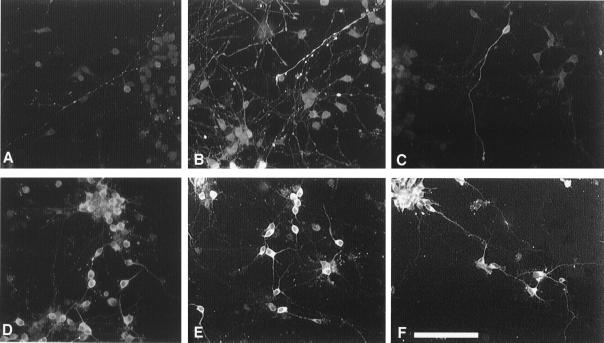
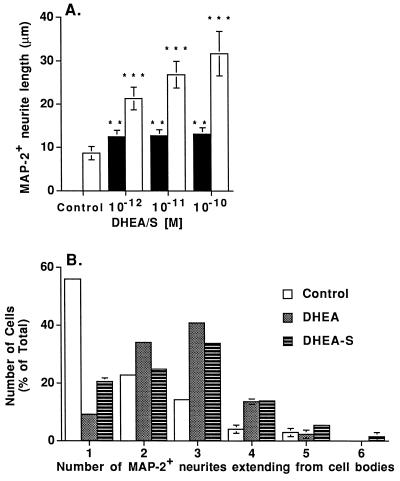
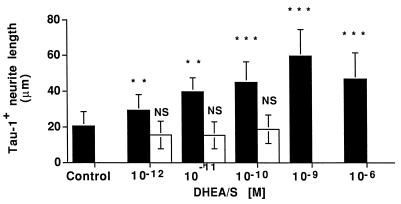
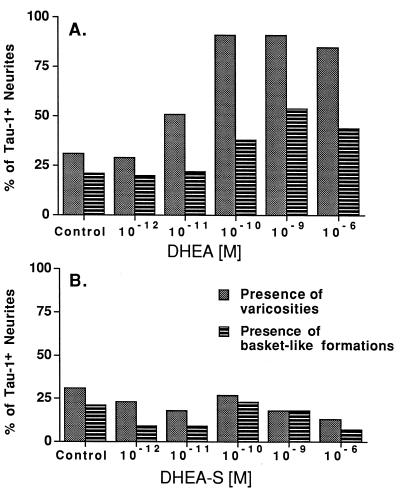
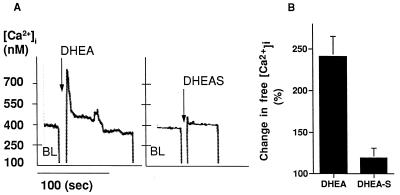
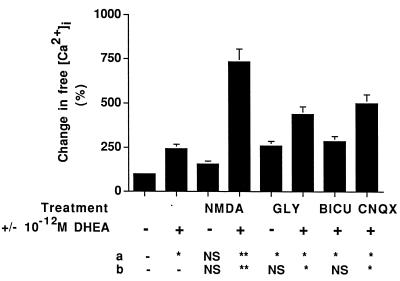
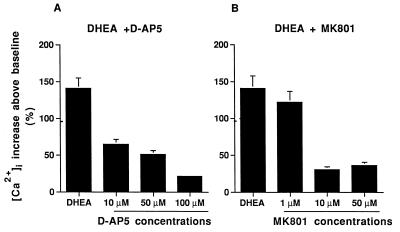
Dehydroepiandrosterone (DHEA) and its sulfate derivative (DHEAS) are the most abundant steroids produced by the human adrenal, but no receptors have been identified for these steroids, and no function for them has been established, other than as precursors for sex steroid synthesis. DHEA and DHEAS are found in brains from many species, and we have shown that enzymes crucial for their synthesis, especially P450c17 (17alpha-hydroxylase/c17,20 lyase), are expressed in a developmentally regulated, region-specific fashion in the developing rodent brain. One region of embryonic expression of P450c17, the neocortical subplate, has been postulated to play a role in guiding cortical projections to their appropriate targets. We therefore determined if products of P450c17 activity, DHEA and DHEAS, regulated the motility and/or growth of neocortical neurons. In primary cultures of mouse embryonic neocortical neurons, DHEA increased the length of neurites containing the axonal marker Tau-1, and the incidence of varicosities and basket-like process formations in a dose-dependent fashion. These effects could be seen at concentrations normally found in the brain. By contrast, DHEAS had no effect on Tau-1 axonal neurites but increased the length of neurites containing the dendritic marker microtubule-associated protein-2. DHEA rapidly increased free intracellular calcium via activation of N-methyl-D-aspartate (NMDA) receptors. These studies provide evidence of mechanisms by which DHEA and DHEAS exert biological actions, show that they have specific functions other than as sex steroid precursors, mediate their effects via non-classic steroid hormone receptors, and suggest that their developmentally regulated synthesis in vivo may play crucial and different roles in organizing the neocortex.
Figures
Comment in
-
Dehydroepiandrosterone (DHEA) and dehydroepiandrosterone sulfate (DHEAS) as neuroactive neurosteroids.Proc Natl Acad Sci U S A. 1998 Apr 14;95(8):4089-91. doi: 10.1073/pnas.95.8.4089. Proc Natl Acad Sci U S A. 1998. PMID: 9539693 Free PMC article. Review. No abstract available.
References
-
- Baulieu E E. Biol Cell. 1991;71:3–10. - PubMed
-
- Mellon S H. J Clin Endocrinol Metab. 1994;78:1003–1008. - PubMed
-
- Le Goascogne C, Robel P, Gouezou M, Sananes N, Baulieu E E, Waterman M. Science. 1987;237:1212–1215. - PubMed
-
- Mellon S H, Deschepper C F. Brain Res. 1993;629:283–292. - PubMed
-
- Dupont E, Simard J, Luu-The V, Labrie F, Pelletier G. Mol Cell Neurosci. 1994;5:119–123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
