

Reversal of age-related alterations in synaptic plasticity by blockade of L-type Ca2+ channels
- PMID: 9547225
- PMCID: PMC6792645
- DOI: 10.1523/JNEUROSCI.18-09-03171.1998
Reversal of age-related alterations in synaptic plasticity by blockade of L-type Ca2+ channels
Abstract
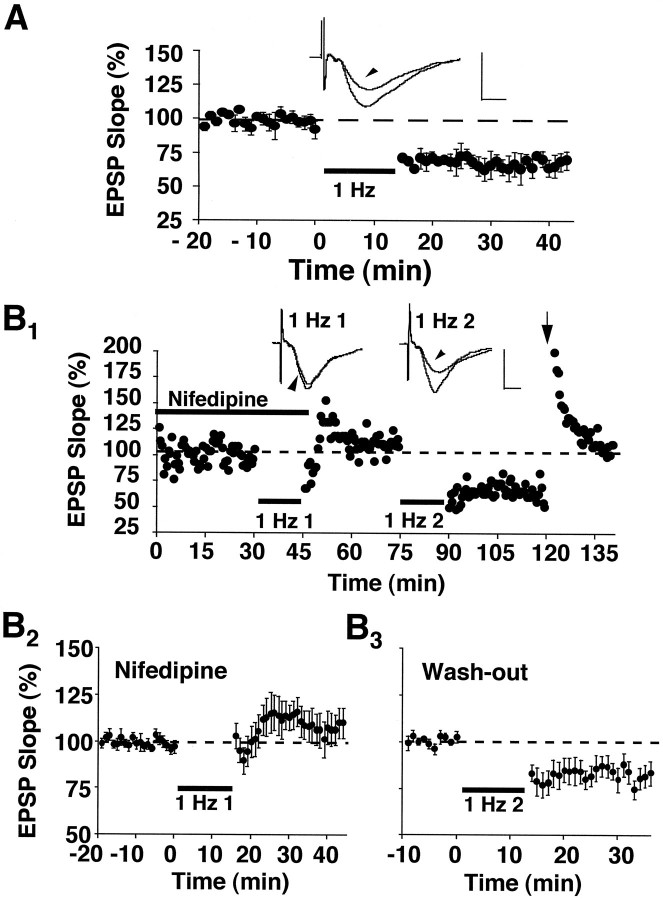
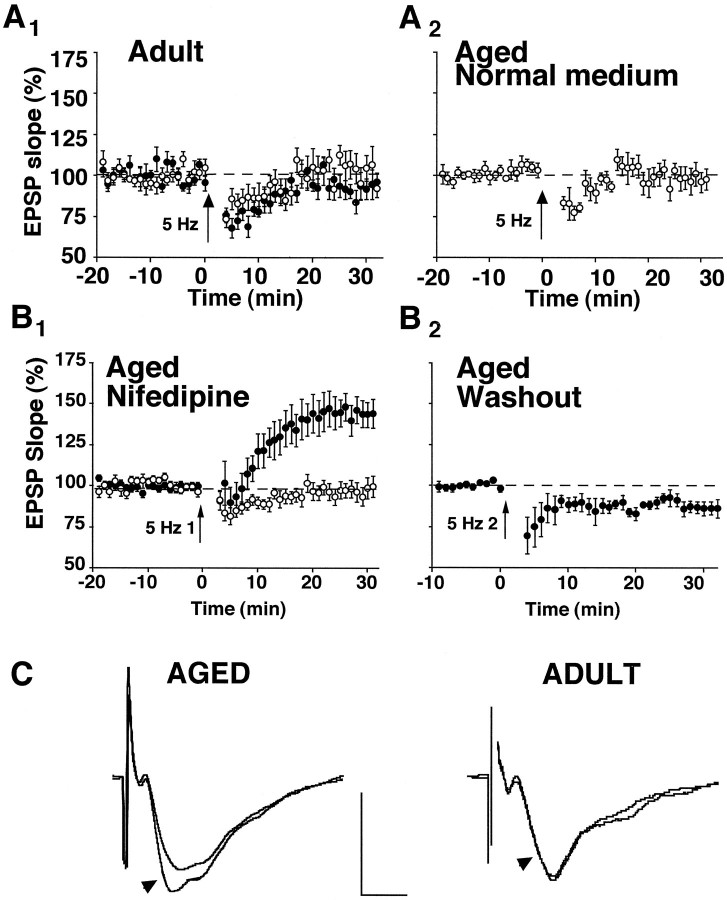
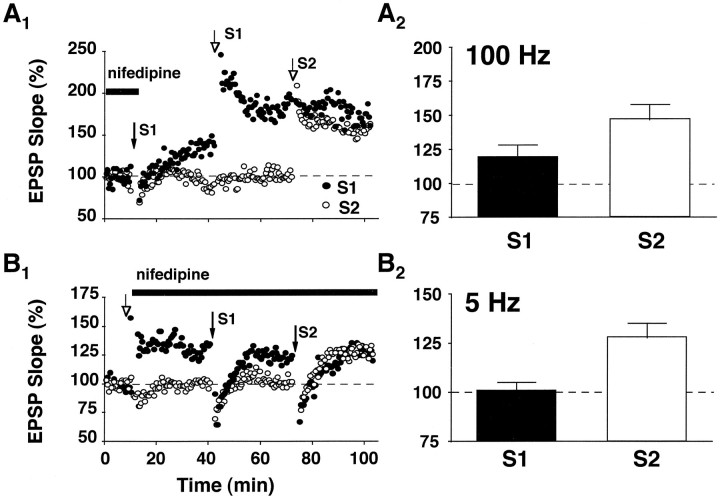
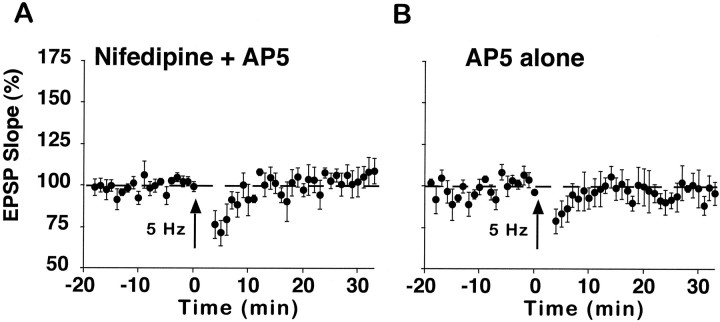
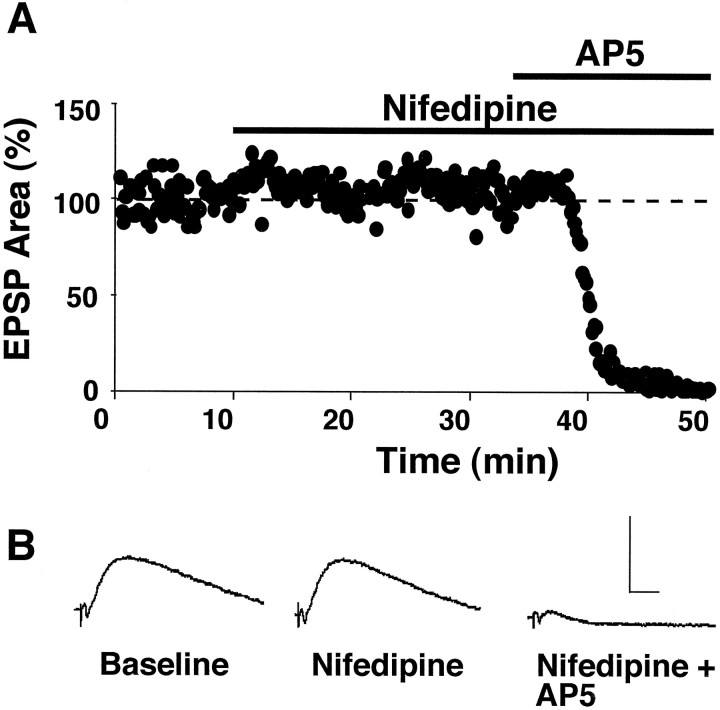
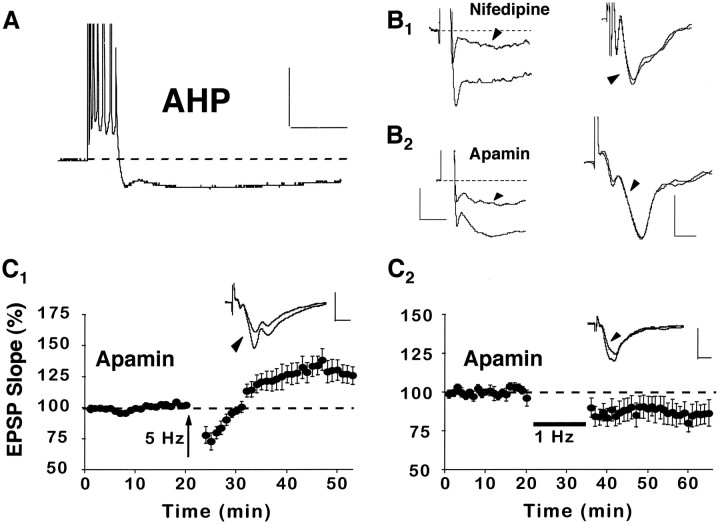
The role of L-type Ca2+ channels in the induction of synaptic plasticity in hippocampal slices of aged (22-24 months) and young adult (4-6 months) male Fischer 344 rats was investigated. Prolonged 1 Hz stimulation (900 pulses) of Schaffer collaterals, which normally depresses CA3/CA1 synaptic strength in aged rat slices, failed to induce long-term depression (LTD) during bath application of the L-channel antagonist nifedipine (10 microM). When 5 Hz stimulation (900 pulses) was used to modify synaptic strength, nifedipine facilitated synaptic enhancement in slices from aged, but not young, adult rats. This enhancement was pathway-specific, reversible, and impaired by the NMDA receptor (NMDAR) antagonist DL-2-amino-5-phosphonopentanoic acid (AP5). Induction of long-term potentiation (LTP) in aged rats, using 100 Hz stimulation, occluded subsequent synaptic enhancement by 5 Hz stimulation, suggesting that nifedipine-facilitated enhancement shares mechanisms in common with conventional LTP. Facilitation of synaptic enhancement by nifedipine likely was attributable to a reduction ( approximately 30%) in the Ca2+-dependent K+-mediated afterhyperpolarization (AHP), because the K+ channel blocker apamin (1 microM) similarly reduced the AHP and promoted synaptic enhancement by 5 Hz stimulation. In contrast, apamin did not block LTD induction using 1 Hz stimulation, suggesting that, in aged rats, the AHP does not influence LTD and LTP induction in a similar way. The results indicate that, during aging, L-channels can (1) facilitate LTD induction during low rates of synaptic activity and (2) impair LTP induction during higher levels of synaptic activation via an increase in the Ca2+-dependent AHP.
Figures
References
-
- Barnes CA, McNaughton BL. Spatial memory and hippocampal synaptic plasticity in senescent and middle-aged rats. In: Stein D, editor. The psychobiology of aging: problems and perspectives. Elsevier; Amsterdam: 1980. pp. 253–272.
-
- Barnes CA, Rao G, Foster TC, McNaughton BL. Region-specific age effects on AMPA sensitivity: electrophysiological evidence for loss of synaptic contacts in hippocampal field CA1. Hippocampus. 1992;2:457–468. - PubMed
-
- Barnes CA, Rao G, McNaughton BL. Functional integrity of NMDA-dependent LTP induction mechanisms across the lifespan of F-344 rats. Learn Memory. 1996;3:124–137. - PubMed
-
- Bear MF. Mechanism for a sliding synaptic modification threshold. Neuron. 1995;15:1–4. - PubMed
-
- Bear MF, Abraham WC. Long-term depression in hippocampus. Annu Rev Neurosci. 1996;19:437–462. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous