

A double-edged kinase Lyn: a positive and negative regulator for antigen receptor-mediated signals
- PMID: 9547345
- PMCID: PMC2212230
- DOI: 10.1084/jem.187.8.1343
A double-edged kinase Lyn: a positive and negative regulator for antigen receptor-mediated signals
Abstract
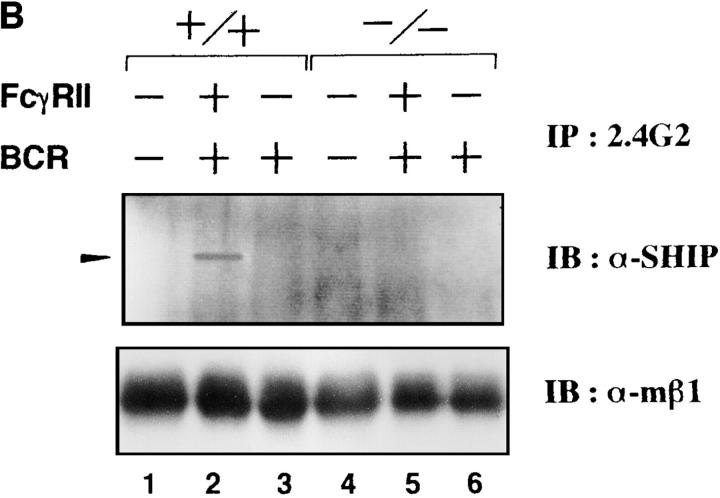
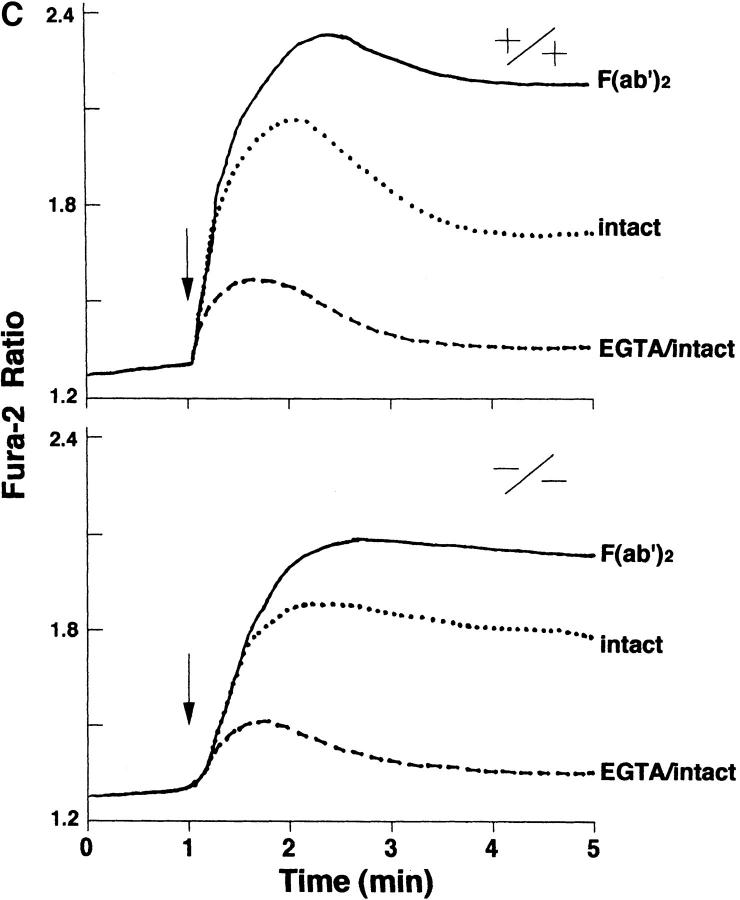
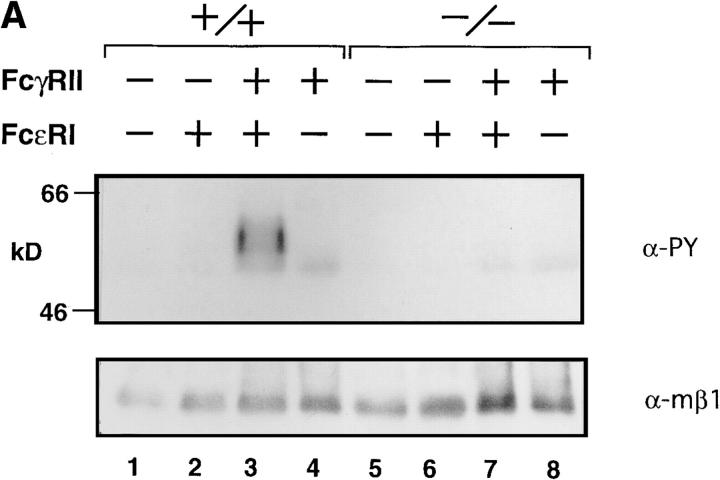
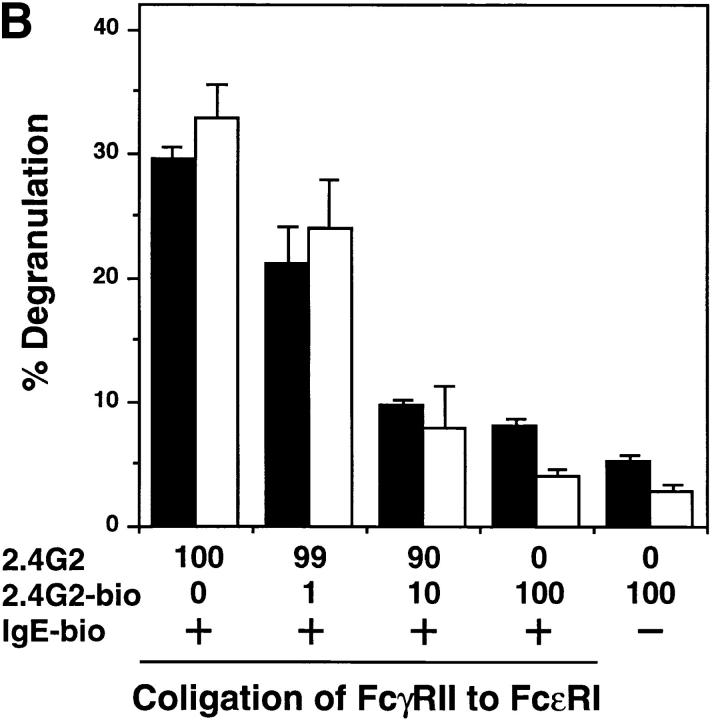
B cells from young lyn-/- mice are hyperresponsive to anti-IgM-induced proliferation, suggesting involvement of Lyn in negative regulation of B cell antigen receptor (BCR)-mediated signaling. Here we show that tyrosine phosphorylation of FcgammaRIIB and CD22 coreceptors, which are important for feedback suppression of BCR-induced signaling, was severely impaired in lyn-/- B cells upon their coligation with the BCR. Hypophosphorylation on tyrosine residues of these molecules resulted in failure of recruiting the tyrosine phosphatase SHP-1 and inositol phosphatase SHIP, SH2-containing potent inhibitors of BCR-induced B cell activation, to the coreceptors. Consequently, lyn-/- B cells exhibited defects in suppressing BCR-induced Ca2+ influx and proliferation. Thus, Lyn is critically important in tyrosine phosphorylation of the coreceptors, which is required for feedback suppression of B cell activation.
Figures
References
-
- Yamanashi Y, Kakiuchi T, Mizuguchi J, Yamamoto T, Toyoshima K. Association of B cell antigen receptor with protein tyrosine kinase Lyn. Science. 1991;251:192–194. - PubMed
-
- Eisenman E, Bolen JB. Engagement of the high-affinity IgE receptor activates src-protein–related tyrosine kinases. Nature. 1992;355:78–80. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
