

Centromere protein B null mice are mitotically and meiotically normal but have lower body and testis weights
- PMID: 9548711
- PMCID: PMC2148459
- DOI: 10.1083/jcb.141.2.309
Centromere protein B null mice are mitotically and meiotically normal but have lower body and testis weights
Abstract
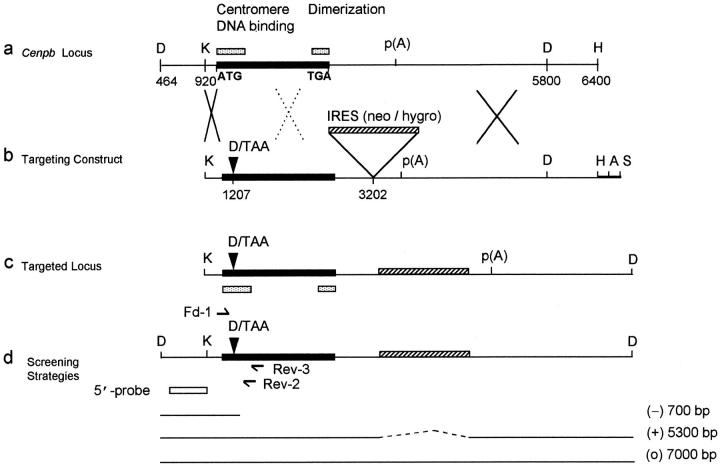
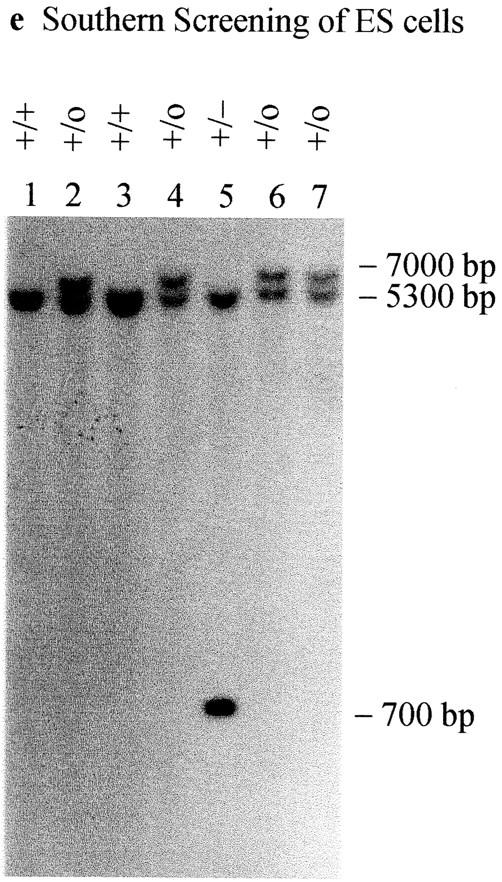
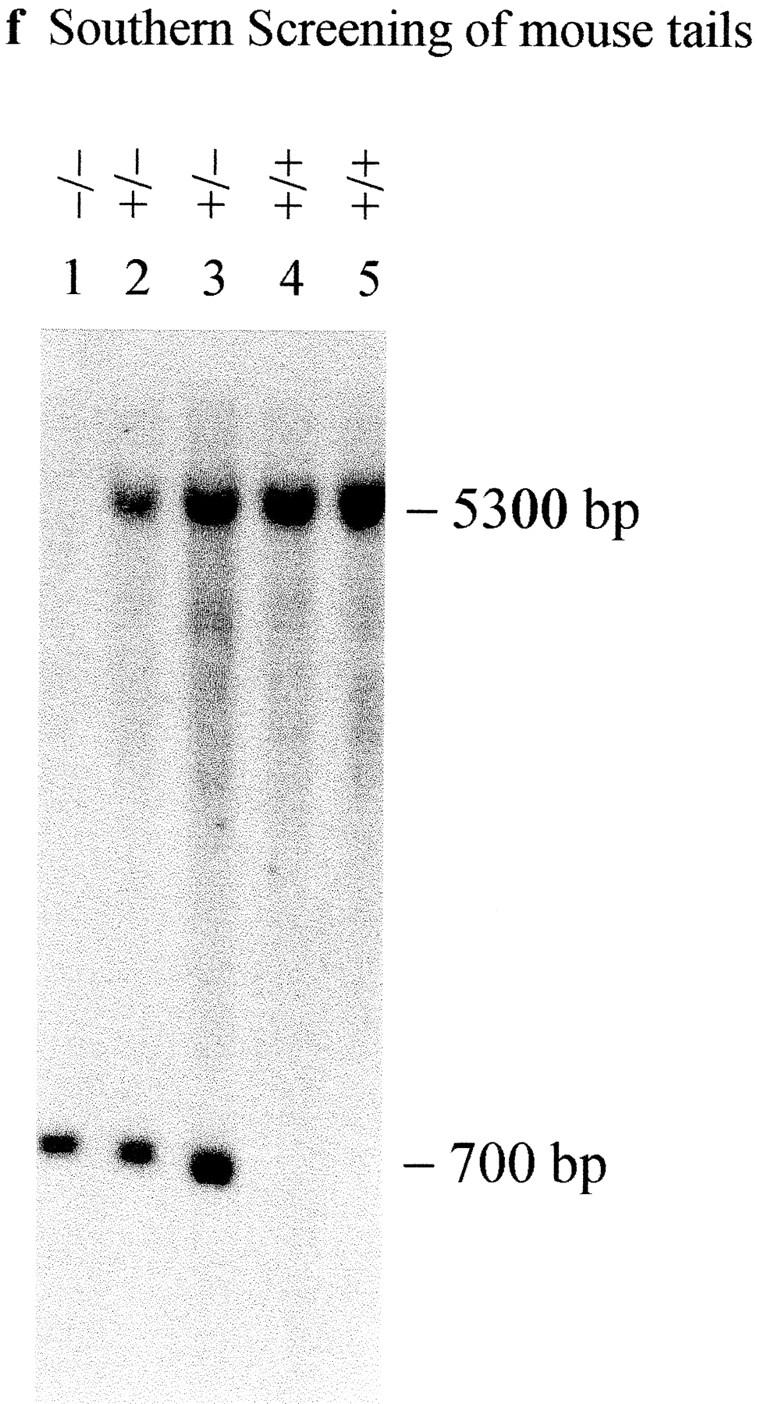
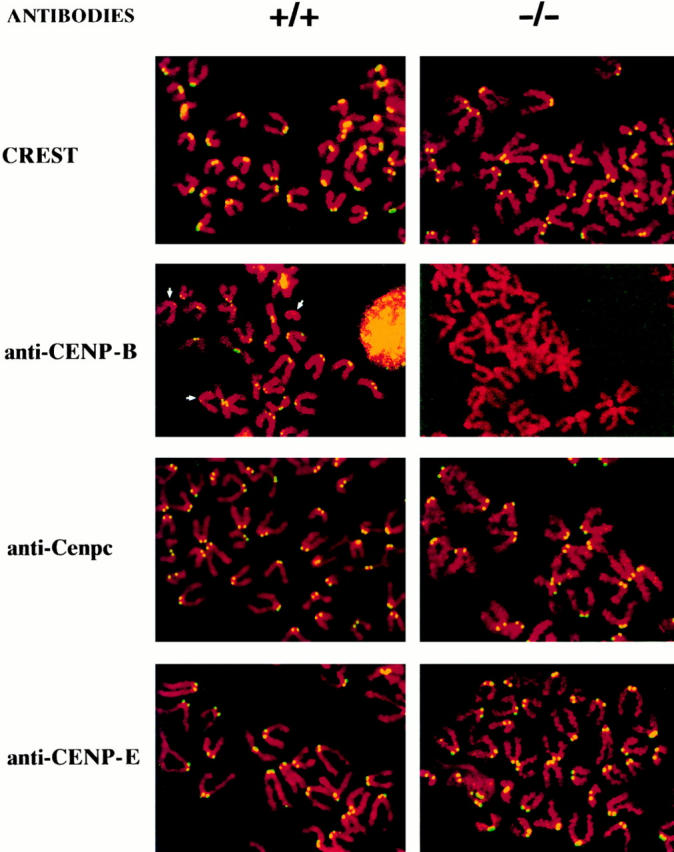
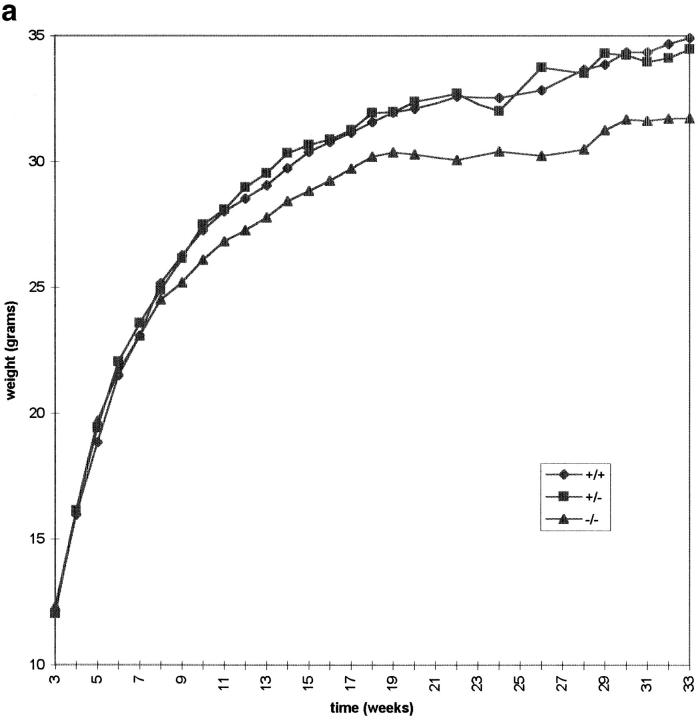
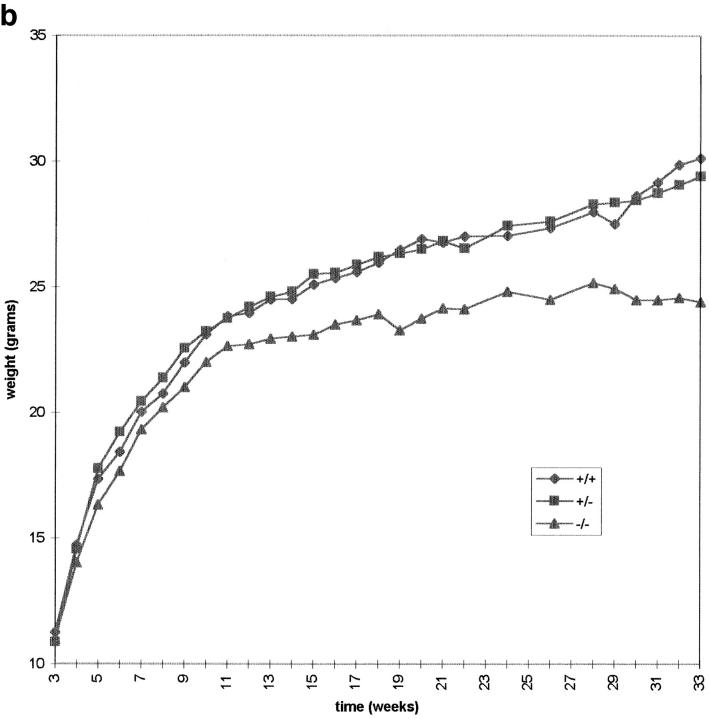
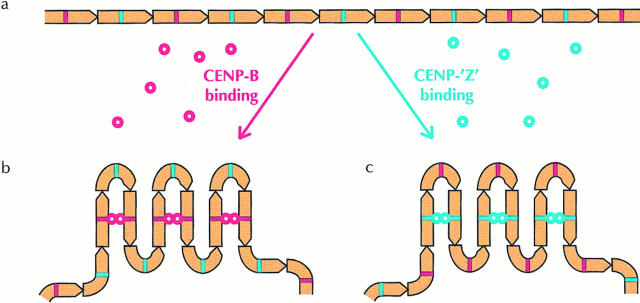
CENP-B is a constitutive centromere DNA-binding protein that is conserved in a number of mammalian species and in yeast. Despite this conservation, earlier cytological and indirect experimental studies have provided conflicting evidence concerning the role of this protein in mitosis. The requirement of this protein in meiosis has also not previously been described. To resolve these uncertainties, we used targeted disruption of the Cenpb gene in mouse to study the functional significance of this protein in mitosis and meiosis. Male and female Cenpb null mice have normal body weights at birth and at weaning, but these subsequently lag behind those of the heterozygous and wild-type animals. The weight and sperm content of the testes of Cenpb null mice are also significantly decreased. Otherwise, the animals appear developmentally and reproductively normal. Cytogenetic fluorescence-activated cell sorting and histological analyses of somatic and germline tissues revealed no abnormality. These results indicate that Cenpb is not essential for mitosis or meiosis, although the observed weight reduction raises the possibility that Cenpb deficiency may subtly affect some aspects of centromere assembly and function, and result in reduced rate of cell cycle progression, efficiency of microtubule capture, and/or chromosome movement. A model for a functional redundancy of this protein is presented.
Figures

References
-
- Bejarano LA, Valdivia MM. Molecular cloning of an intronless gene for the hamster centromere antigen CENP-B. Biochim Biophys Acta. 1996;1307:21–25. - PubMed
-
- Bernat RL, Delannoy MR, Rothfield NF, Earnshaw WC. Disruption of centromere assembly during interphase inhibits kinetochore morphogenesis and function in mitosis. Cell. 1991;66:1229–1238. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
