

Rho-mediated contractility exposes a cryptic site in fibronectin and induces fibronectin matrix assembly
- PMID: 9548730
- PMCID: PMC2148448
- DOI: 10.1083/jcb.141.2.539
Rho-mediated contractility exposes a cryptic site in fibronectin and induces fibronectin matrix assembly
Abstract
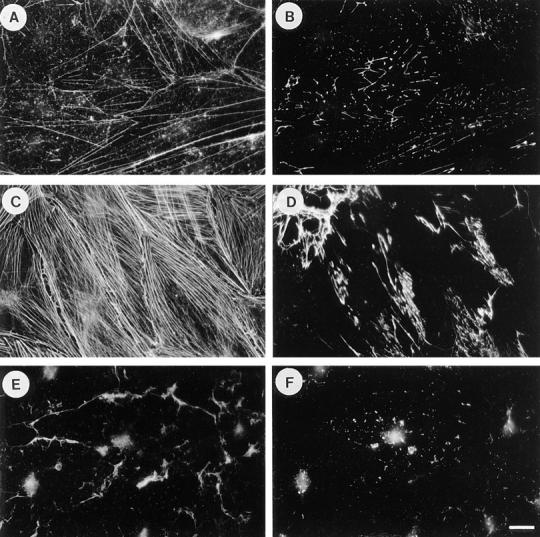
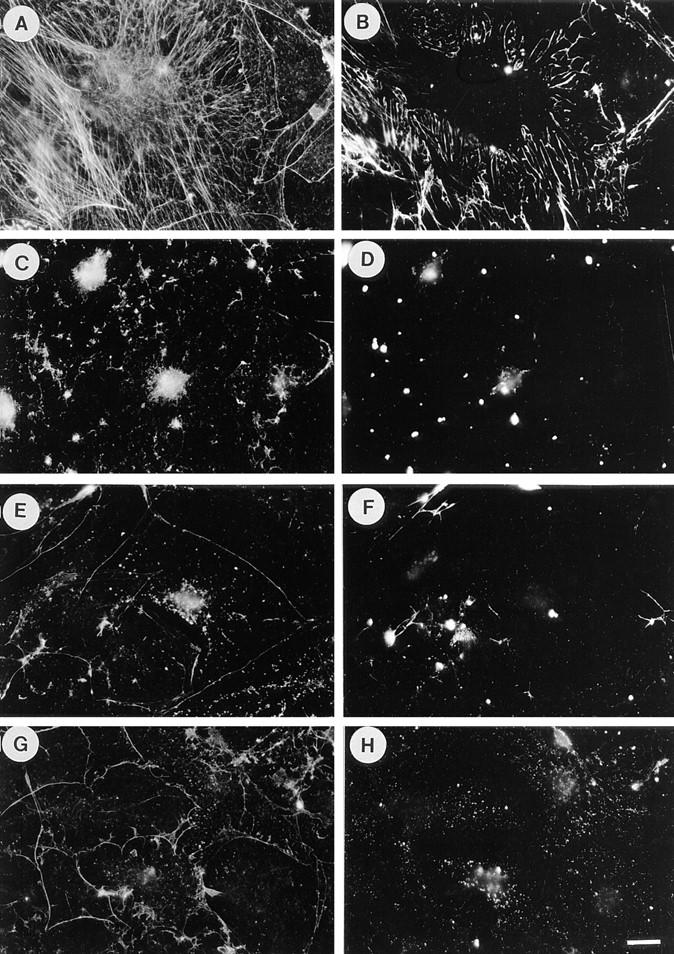
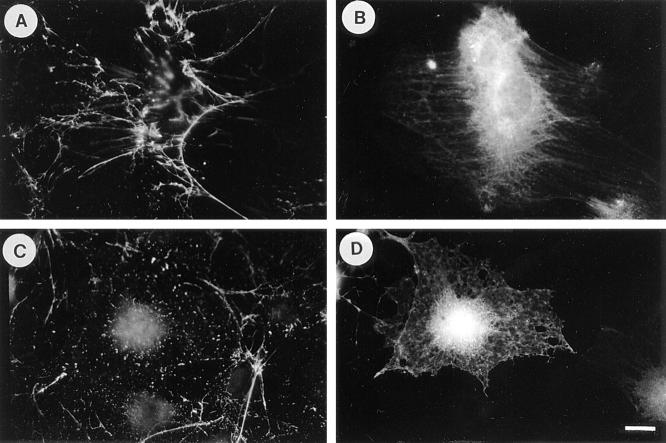
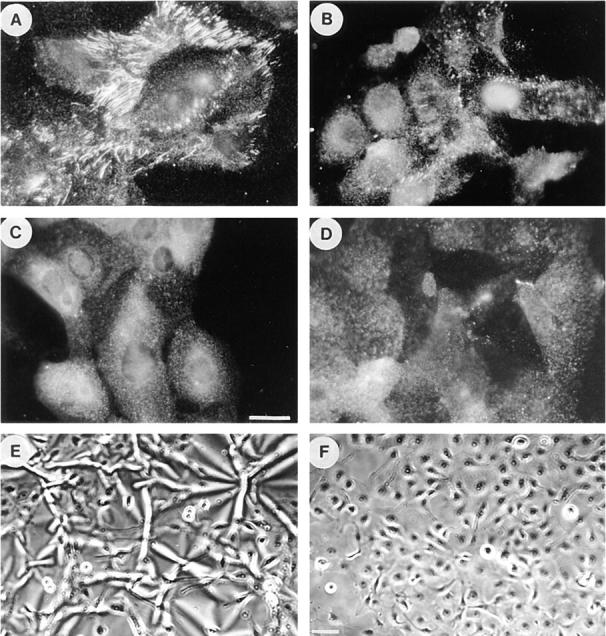
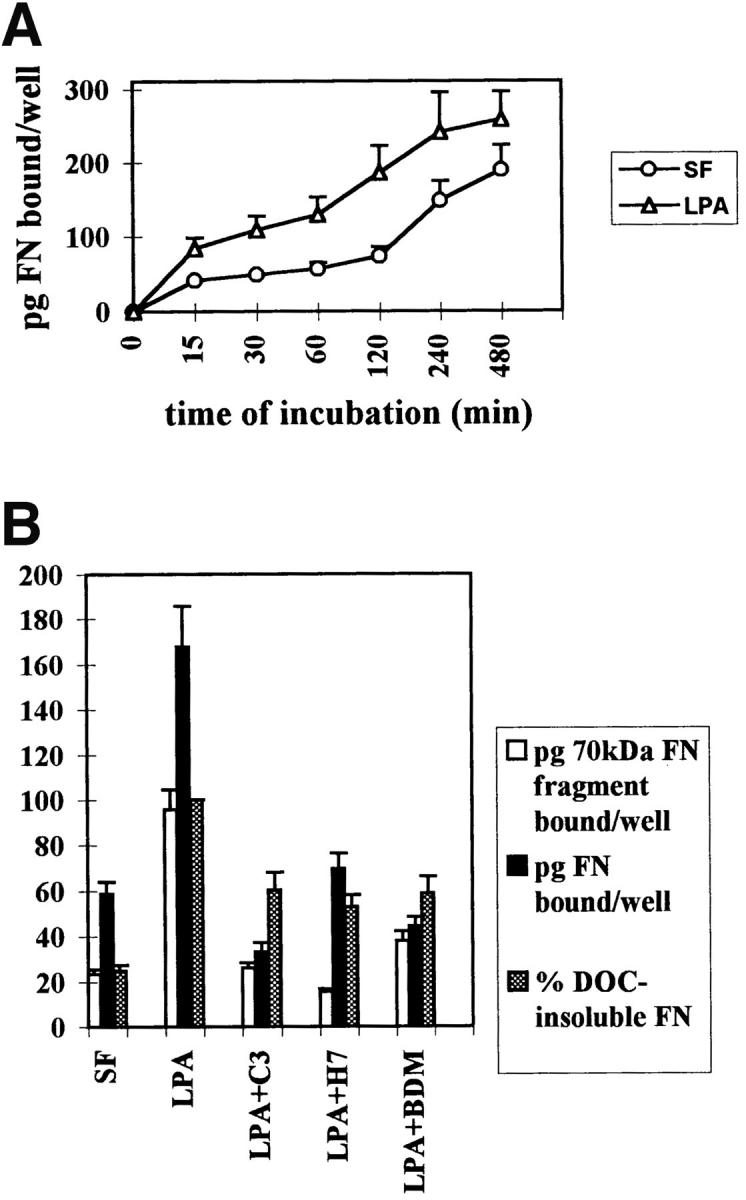
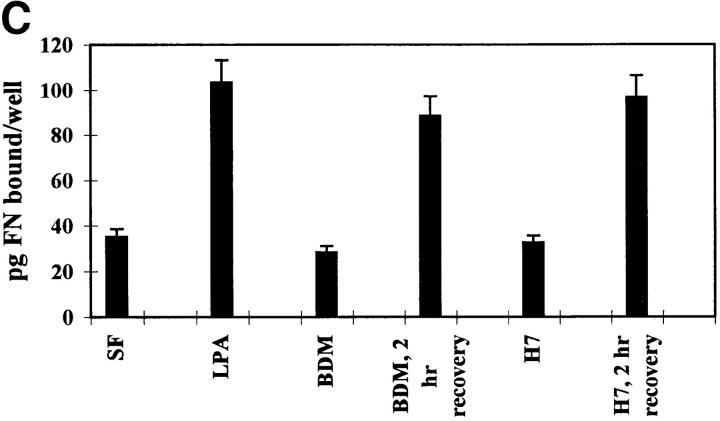
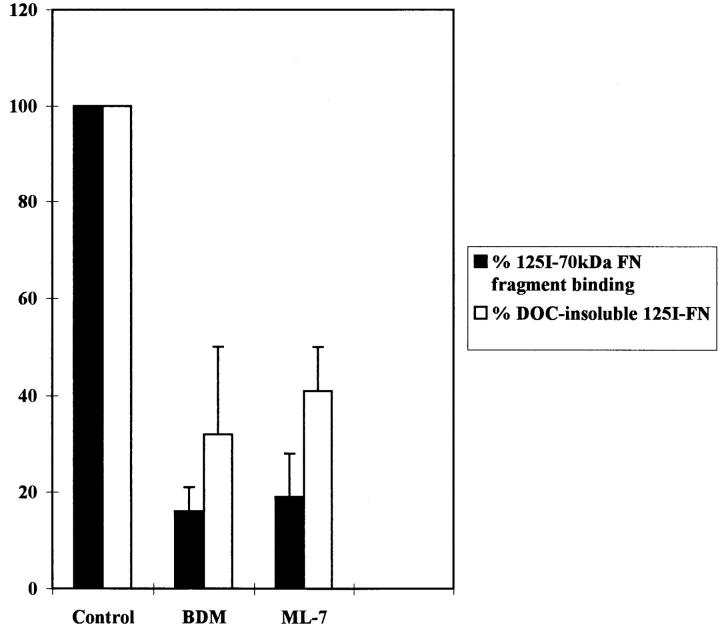
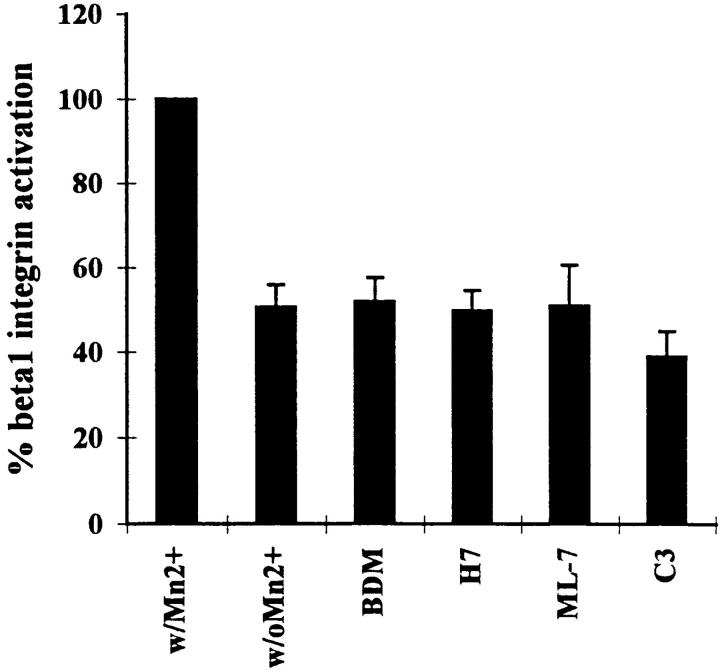
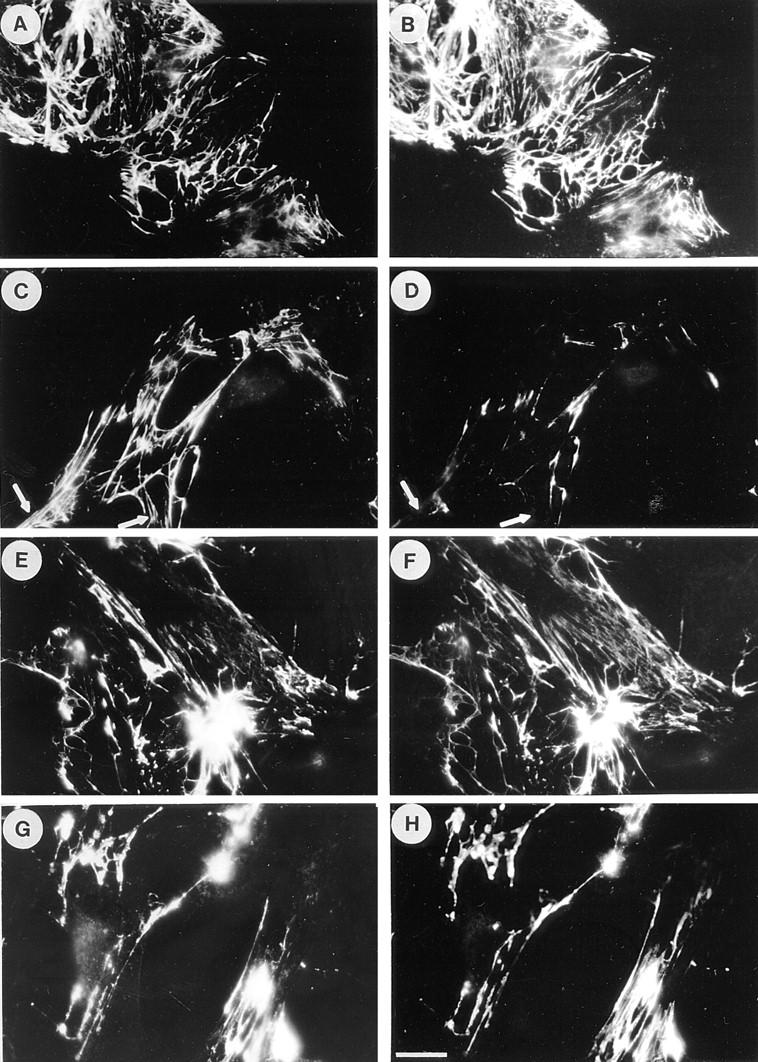
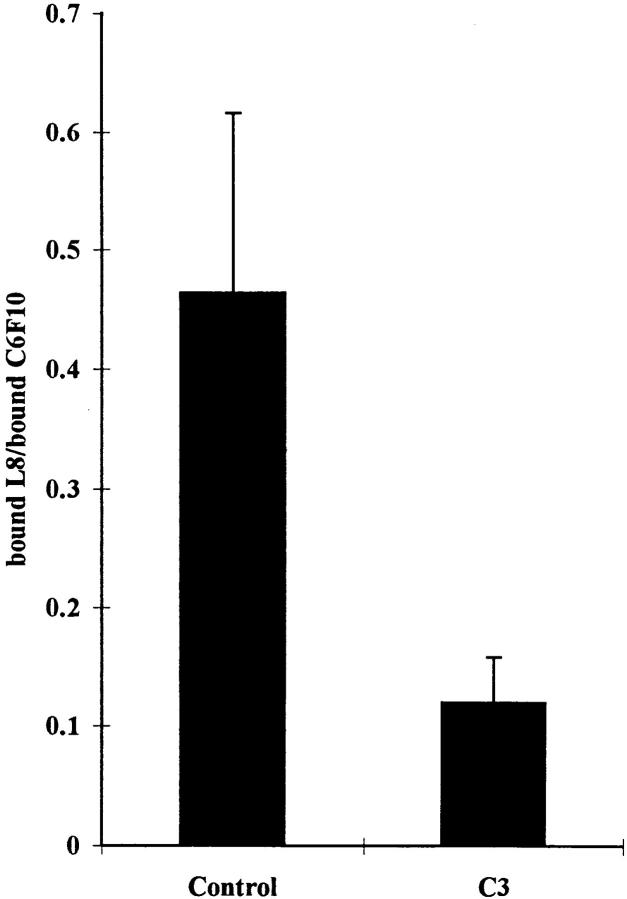
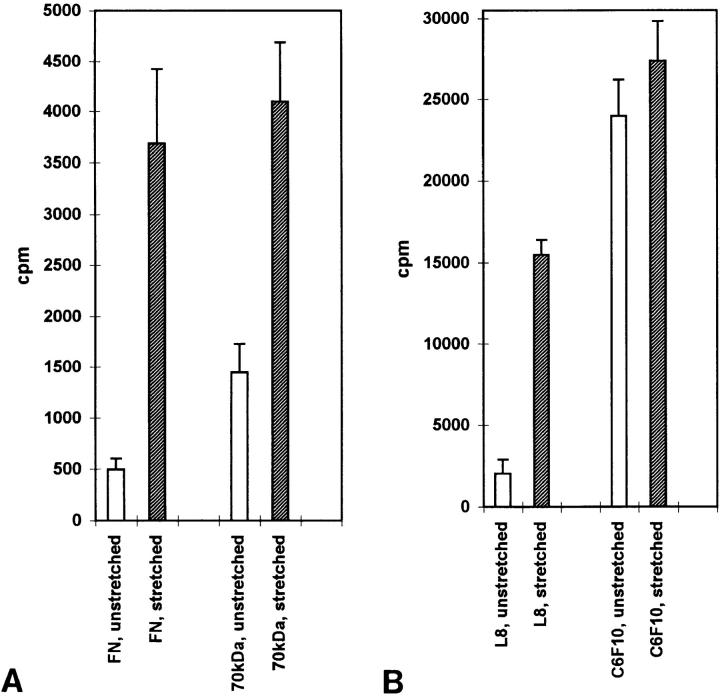
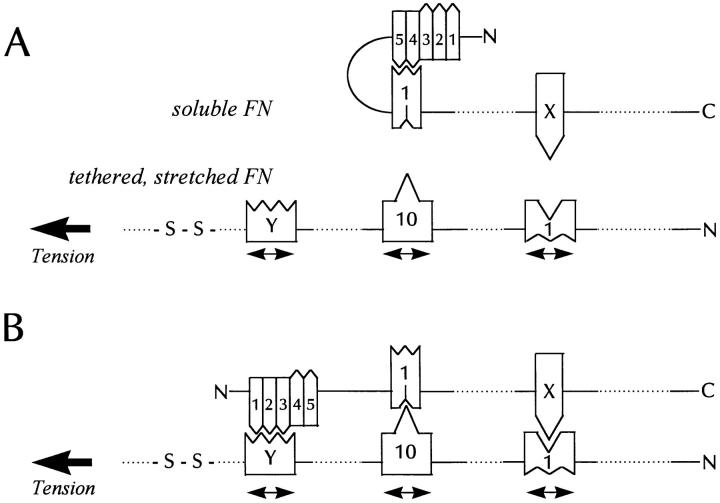
Many factors influence the assembly of fibronectin into an insoluble fibrillar extracellular matrix. Previous work demonstrated that one component in serum that promotes the assembly of fibronectin is lysophosphatidic acid (Zhang, Q., W.J. Checovich, D.M. Peters, R.M. Albrecht, and D.F. Mosher. 1994. J. Cell Biol. 127:1447-1459). Here we show that C3 transferase, an inhibitor of the low molecular weight GTP-binding protein Rho, blocks the binding of fibronectin and the 70-kD NH2-terminal fibronectin fragment to cells and blocks the assembly of fibronectin into matrix induced by serum or lysophosphatidic acid. Microinjection of recombinant, constitutively active Rho into quiescent Swiss 3T3 cells promotes fibronectin matrix assembly by the injected cells. Investigating the mechanism by which Rho promotes fibronectin polymerization, we have used C3 to determine whether integrin activation is involved. Under conditions where C3 decreases fibronectin assembly we have only detected small changes in the state of integrin activation. However, several inhibitors of cellular contractility, that differ in their mode of action, inhibit cell binding of fibronectin and the 70-kD NH2-terminal fibronectin fragment, decrease fibronectin incorporation into the deoxycholate insoluble matrix, and prevent fibronectin's assembly into fibrils on the cell surface. Because Rho stimulates contractility, these results suggest that Rho-mediated contractility promotes assembly of fibronectin into a fibrillar matrix. One mechanism by which contractility could enhance fibronectin assembly is by tension exposing cryptic self-assembly sites within fibronectin that is being stretched. Exploring this possibility, we have found a monoclonal antibody, L8, that stains fibronectin matrices differentially depending on the state of cell contractility. L8 was previously shown to inhibit fibronectin matrix assembly (Chernousov, M.A., A.I. Faerman, M.G. Frid, O.Y. Printseva, and V.E. Koteliansky. 1987. FEBS (Fed. Eur. Biochem. Soc.) Lett. 217:124-128). When it is used to stain normal cultures that are developing tension, it reveals a matrix indistinguishable from that revealed by polyclonal anti-fibronectin antibodies. However, the staining of fibronectin matrices by L8 is reduced relative to the polyclonal antibody when the contractility of cells is inhibited by C3. We have investigated the consequences of mechanically stretching fibronectin in the absence of cells. Applying a 30-35% stretch to immobilized fibronectin induced binding of soluble fibronectin, 70-kD fibronectin fragment, and L8 monoclonal antibody. Together, these results provide evidence that self-assembly sites within fibronectin are exposed by tension.
Figures
References
-
- Aguirre KM, McCormick RJ, Schwarzbauer JE. Fibronectin self-association is mediated by complementary sites within the amino-terminal one-third of the molecule. J Biol Chem. 1994;269:27863–27868. - PubMed
-
- Ali IU, Hynes RO. Effects of cytochalasin B and colchicine on attachment of a major surface protein of fibroblasts. Biochim Biophys Acta. 1977;471:16–24. - PubMed
-
- Avnur Z, Geiger B. The removal of extracellular fibronectin from areas of cell–substrate contact. Cell. 1981;25:121–132. - PubMed
-
- Banes AJ, Link GW, Gilbert JA, Tran R Son Tay, and O. Monbureau. Culturing cells in a mechanically active environment. Am Biotechnol Lab. 1990;8:12–22. - PubMed
-
- Blystone SD, Kaplan JE. Isolation of an amino-terminal fibronectin-binding protein on human U937 cells and rat peritoneal macrophages. J Biol Chem. 1992;267:3968–3975. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
