

doi: 10.1101/gad.12.8.1092.
A specific, nonproliferative role for E2F-5 in choroid plexus function revealed by gene targeting
Affiliations
- PMID: 9553039
- PMCID: PMC316727
- DOI: 10.1101/gad.12.8.1092
Item in Clipboard
A specific, nonproliferative role for E2F-5 in choroid plexus function revealed by gene targeting
Genes Dev.
.
Abstract
Homozygous E2F-5 knockout embryos and mice have been generated. Although embryonic development appeared normal, newborn mice developed nonobstructive hydrocephalus, suggesting excessive cerebrospinal fluid (CSF) production. Although the CSF-producing choroid plexus displayed normal cellular organization, it contained abundant electron-lucent epithelial cells, consistent with excessive CSF secretory activity. Moreover, E2F-5 CNS expression in normal animals was largely confined to the choroid plexus. Cell cycle kinetics were not perturbed in homozygous knockout embryo fibroblasts. Thus, E2F-5 is not essential for cell proliferation. Rather, it affects the secretory behavior of a differentiated neural tissue.
Figures
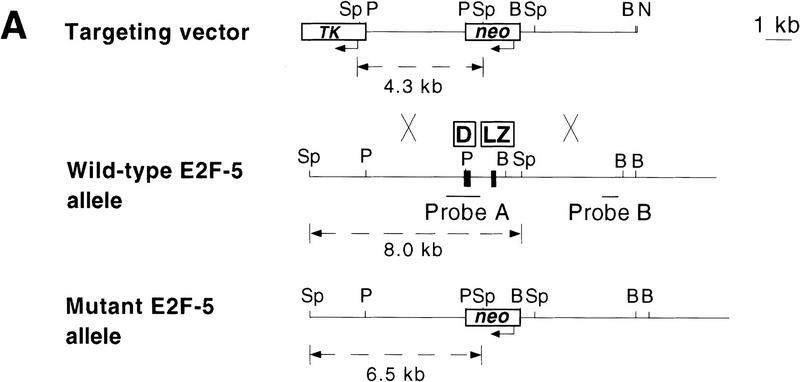
Targeting strategy, germ-line transmission, and gene expression analysis. (A) Organization of the targeting vector, mouse E2F-5 gene, and the allele resulting from homologous recombination. Coding regions from the DNA-binding (D) and leucine zipper (LZ) domains are boxed. A 1.2-kb XbaI fragment (probe A) was used to screen for recombinant alleles, and the sizes of recombinant, wild-type, or randomly integrated transgene fragments are shown following SphI digestion (broken line). A 0.8-kb EcoRI fragment (probe B) and neo cDNA were used for confirmatory screening. (TK) Thymidine kinase; (neo) neomycin; (Sp) SphI; (P) PstI; (B) BamHI; (N) NotI. (B) Southern blot analysis of SphI-digested tail DNA from a litter of F2 mice confirming germ-line transmission using probe A. (C) Northern blot analysis of total mRNA from ES cell clones and brains of neonatal F2 mice. Genotypes are indicated.
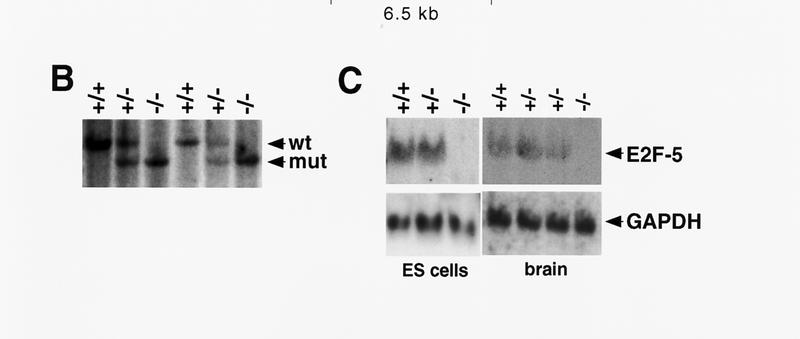
Targeting strategy, germ-line transmission, and gene expression analysis. (A) Organization of the targeting vector, mouse E2F-5 gene, and the allele resulting from homologous recombination. Coding regions from the DNA-binding (D) and leucine zipper (LZ) domains are boxed. A 1.2-kb XbaI fragment (probe A) was used to screen for recombinant alleles, and the sizes of recombinant, wild-type, or randomly integrated transgene fragments are shown following SphI digestion (broken line). A 0.8-kb EcoRI fragment (probe B) and neo cDNA were used for confirmatory screening. (TK) Thymidine kinase; (neo) neomycin; (Sp) SphI; (P) PstI; (B) BamHI; (N) NotI. (B) Southern blot analysis of SphI-digested tail DNA from a litter of F2 mice confirming germ-line transmission using probe A. (C) Northern blot analysis of total mRNA from ES cell clones and brains of neonatal F2 mice. Genotypes are indicated.
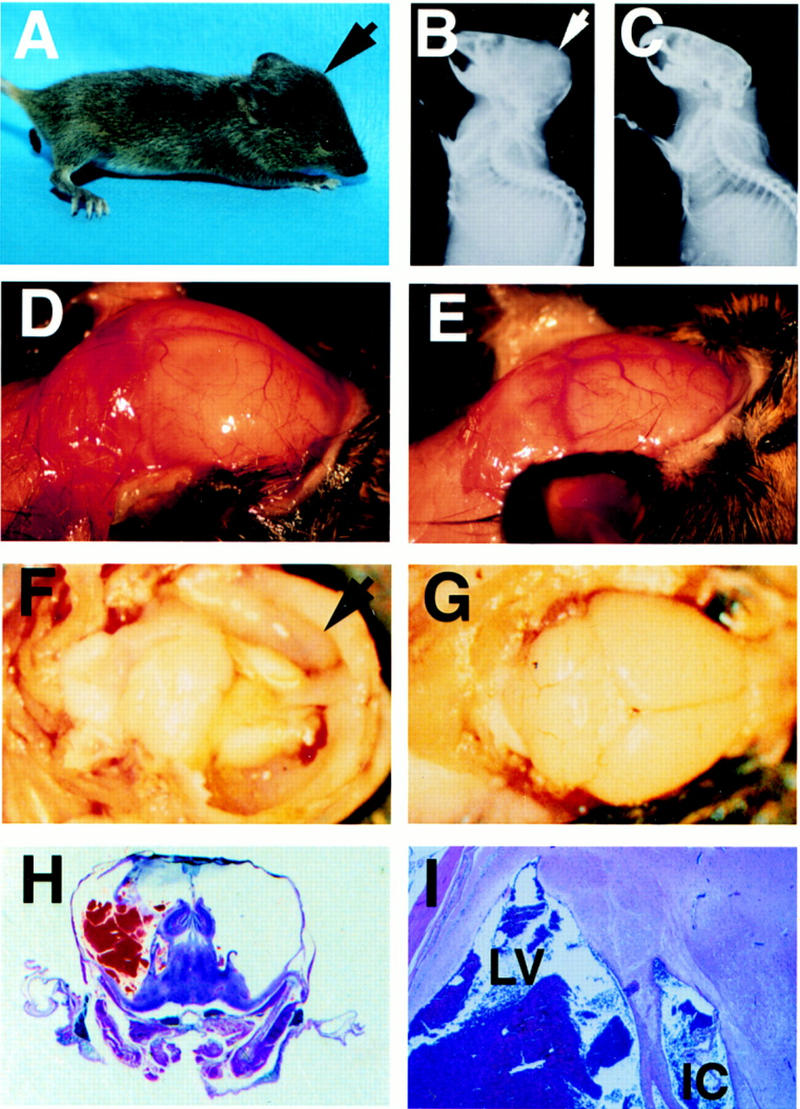
E2F-5−/− mice develop hydrocephalus. (A) Profile of knockout mouse with domed head (arrow). Expanded calvarium (arrow) in homozygous knockout mouse detected by X-radiography (B) compared to a wild-type littermate (C). Expanded calvarium and underlying brain seen in knockout (D) compared to healthy littermate (E). (F) Thin rim of expanded cerebral cortex (anterior cortex arrowed) overlying dilated lateral ventricles; visible underlying structures include (from right to left) hippocampus, thalamus, and cerebellum. (G) Cerebrum of normal, smaller littermate; parietal lobes (right) and cerebellum (left) are visible. (H) H&E-stained coronal section of the cranium revealing massive ventricular dilation, thinning of surrounding neural structures, an intraventricular thrombus, and compression of the hippocampus (superior structures) and underlying thalamus. (I) Dilated lateral ventricle (LV) containing thrombus and intracerebral hemorrhage (IC). (J,L,N) Cerebral cortex of E2F-5−/− pups compared to (K,M,O) the same tissue from a wild-type littermate at D1 (J,K), D4 (L,M), and D7 (N,O). No ventricular dilation was seen at D1, but was apparent by D4. (P,Q), Alcian blue (cartilage) and alizarin red (bone) staining of cleared E16 and E19 E2F-5−/− embryos, respectively. The interparietal bone (i, arrow), supraoccipital bone (s, arrow) and humerus (h, arrow) were normally developed compared to those of wild-type littermates (not shown).
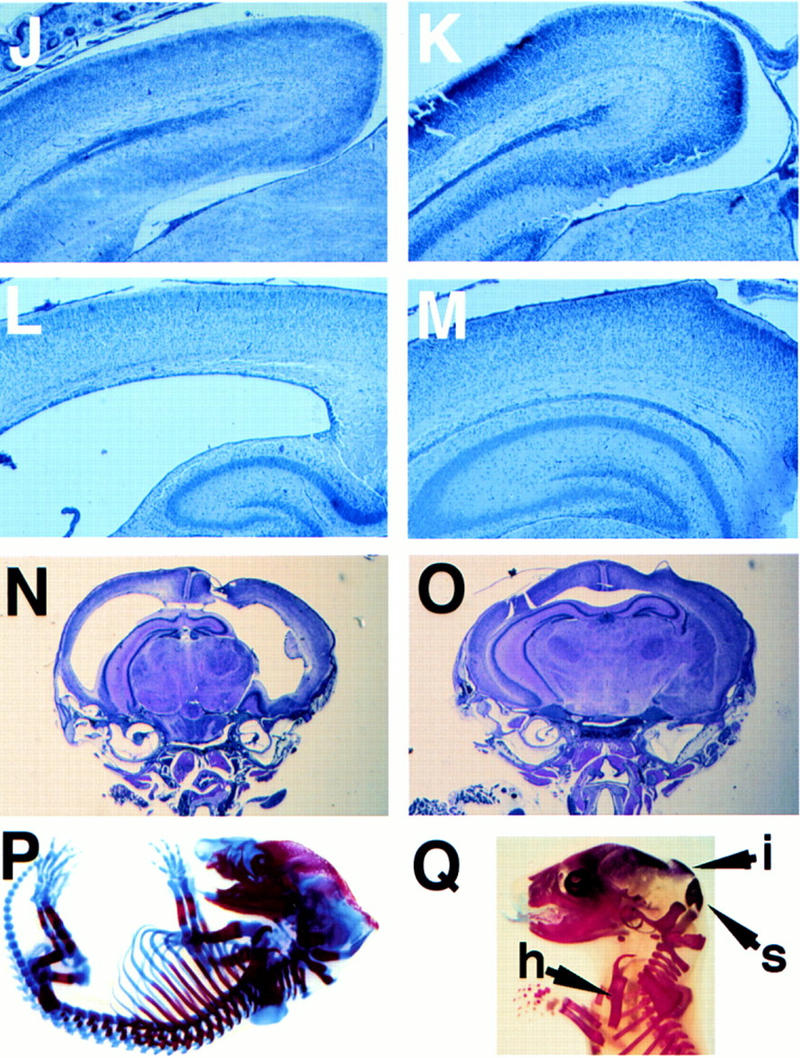
E2F-5−/− mice develop hydrocephalus. (A) Profile of knockout mouse with domed head (arrow). Expanded calvarium (arrow) in homozygous knockout mouse detected by X-radiography (B) compared to a wild-type littermate (C). Expanded calvarium and underlying brain seen in knockout (D) compared to healthy littermate (E). (F) Thin rim of expanded cerebral cortex (anterior cortex arrowed) overlying dilated lateral ventricles; visible underlying structures include (from right to left) hippocampus, thalamus, and cerebellum. (G) Cerebrum of normal, smaller littermate; parietal lobes (right) and cerebellum (left) are visible. (H) H&E-stained coronal section of the cranium revealing massive ventricular dilation, thinning of surrounding neural structures, an intraventricular thrombus, and compression of the hippocampus (superior structures) and underlying thalamus. (I) Dilated lateral ventricle (LV) containing thrombus and intracerebral hemorrhage (IC). (J,L,N) Cerebral cortex of E2F-5−/− pups compared to (K,M,O) the same tissue from a wild-type littermate at D1 (J,K), D4 (L,M), and D7 (N,O). No ventricular dilation was seen at D1, but was apparent by D4. (P,Q), Alcian blue (cartilage) and alizarin red (bone) staining of cleared E16 and E19 E2F-5−/− embryos, respectively. The interparietal bone (i, arrow), supraoccipital bone (s, arrow) and humerus (h, arrow) were normally developed compared to those of wild-type littermates (not shown).

Lethality from hydrocephalus, MEF proliferation, and cell cycle analysis. (A) Median survival rate for E2F-5−/− mice (strain 33). Animals were sacrificed for autopsy when terminally ill. (B) Cell cycle position of asynchronous wild-type (n = 5) and knockout (n = 4) MEFs determined by BrdU incorporation and FACS analysis. (C) Cumulative growth of primary E2F-5 MEFs. Primary fibroblasts (passage 0) were seeded on 100-mm-diam. plates and sequentially transferred to three plates, each, at 106 cells/plate every 2–3 days. Cell growth was determined after each passage. Cumulative growth was determined and plotted against the number of days of passage. (D) BrdU incorporation of MEFs following synchronization and serum release. Genotypes are indicated.
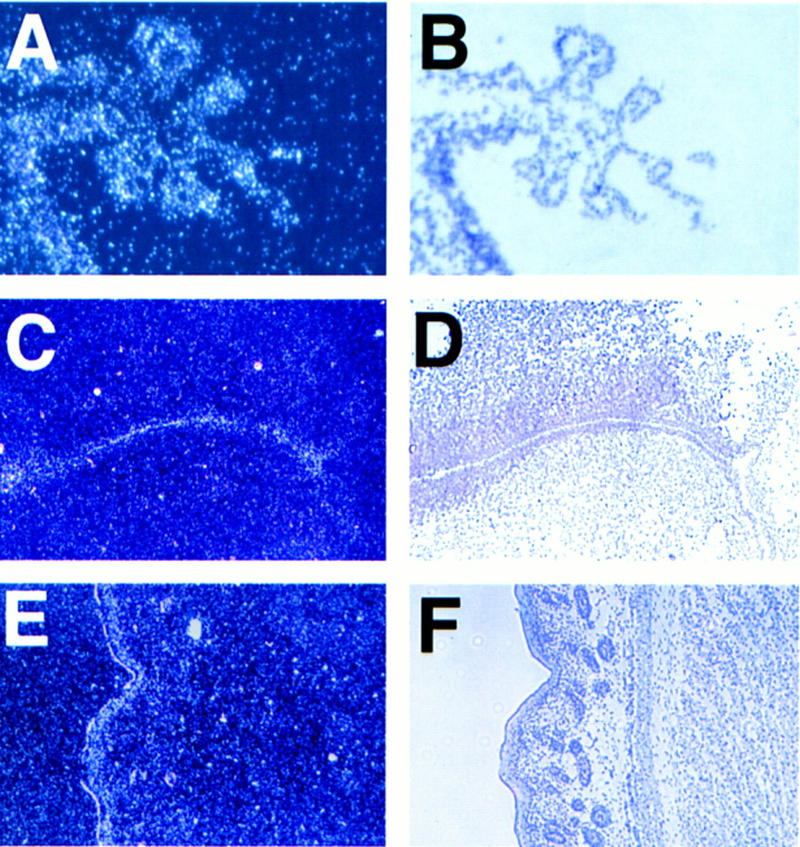
E2F-5 is expressed in the choroid plexus epithelium and periventricular region in newborn mice. Dark-field (A,C,E) and corresponding light-field (B,D,F) micrographs hybridized to an E2F-5 antisense probe. The relevant tissues were D1 choroid plexus (A,B) derived from fourth ventricle (sagittal section); D1 cortex (C,D) showing positive grains in the ependymal cells lining the lateral ventricles (coronal section); D3 skin (E,F) with positive grains in the suprabasal epidermis (sagittal section). A control, sense probe did not hybridize to these tissues (data not shown).

Light microscopic and ultrastructural analysis of E2F-5 mutant choroid plexus. Choroid plexus within the dilated lateral ventricle of a homozygous knockout animal showing normal morphology by light microscopy (A) compared to choroid plexus (arrow) from a wild-type littermate (B). (C)E2F-5−/− choroid plexus showing numerous electron lucent (light) cells, one of which is arrowed. The microvillus border is irregular and disorganized (original magnification 2940×). (D) In contrast, a wild-type littermate shows uniform cellular electron density, apart from an occasional light cell (2940×). (E) Magnified view showing light cells in E2F-5−/− choroid plexus. Ultrastructural organelles appear intact (4750×). EM sections were from 4-day-old neonates.
References
-
- Adams PD, Kaelin WG., Jr The cellular effects of E2F overexpression. Curr Top Microbiol Immunol. 1996;208:79–93. - PubMed
-
- Buck V, Allen KE, Sørensen T, Bybee A, Hijmans EM, Voorhoeve PM, La Thangue N. Molecular and functional characterisation of E2F-5, a new member of the E2F family. Oncogene. 1995;11:31–38. - PubMed
-
- Cobrinik D. Regulatory interactions among E2Fs and cell cycle control proteins. Curr Top Microbiol Immunol. 1996;208:31–61. - PubMed
-
- Cobrinik D, Whyte P, Peeper D, Jacks T, Weinberg RA. Cell cycle-specific association of E2F with the p130 E1A-binding protein. Genes & Dev. 1993;7:2392–2404. - PubMed
-
- Cobrinik D, Lee M-H, Hannon G, Mulligan G, Bronson RT, Dyson N, Harlow E, Beach D, Weinberg RA, Jacks T. Shared role of the pRB-related p130 and p107 proteins in limb development. Genes & Dev. 1996;10:1633–1644. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases