

Coding sequences enhance internal initiation of translation by hepatitis A virus RNA in vitro
- PMID: 9557637
- PMCID: PMC109577
- DOI: 10.1128/JVI.72.5.3571-3577.1998
Coding sequences enhance internal initiation of translation by hepatitis A virus RNA in vitro
Abstract
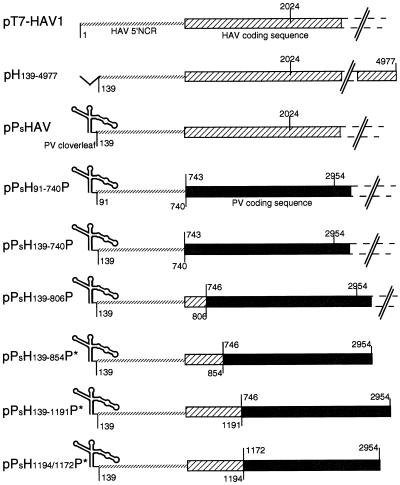
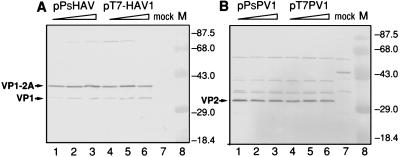
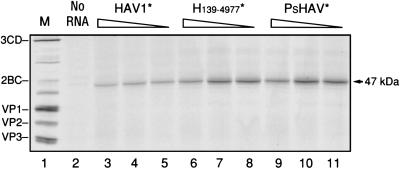
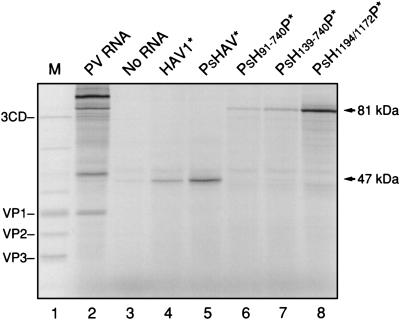
Hepatitis A virus (HAV), unlike other picornaviruses, has a slow-growth phenotype in permissive cell lines and in general does not induce host cell cytopathology. Although there are no published reports of productive infection of HeLa cells by HAV, HAV RNA appears to be readily translated in HeLa cells when transcribed by T7 RNA polymerase provided by a recombinant vaccinia virus. The 5' noncoding region of HAV was fused to poliovirus (PV) coding sequences to determine the effect on translation efficiency in HeLa cell extracts in vitro. Conditions were optimized for utilization of the HAV internal ribosome entry segment (IRES). Transcripts from chimeric constructs fused precisely at the initiation codon were translated very poorly. However, chimeric RNAs which included 114 or more nucleotides from the HAV capsid coding sequences downstream of the initiation codon were translated much more efficiently than those lacking these sequences, making HAV-directed translation efficiency similar to that directed by the PV IRES. Sixty-six nucleotides were insufficient to confer increased translation efficiency. The most 5'-terminal HAV 138 nucleotides, previously determined to be upstream of the IRES, had an inhibitory effect on translation efficiency. Constructs lacking these terminal sequences, or those in which the PV 5'-terminal sequences replaced those from HAV, translated three- to fourfold better than those with the intact HAV 5'-terminal end.
Figures
References
-
- Andino R, Silvera D, Suggett S D, Achacoso P L, Miller C J, Baltimore D, Feinberg M B. Engineering poliovirus as a vaccine vector for the expression of diverse antigens. Science. 1994;265:1448–1451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
