

Foot-and-mouth disease virus virulent for cattle utilizes the integrin alpha(v)beta3 as its receptor
- PMID: 9557639
- PMCID: PMC109579
- DOI: 10.1128/JVI.72.5.3587-3594.1998
Foot-and-mouth disease virus virulent for cattle utilizes the integrin alpha(v)beta3 as its receptor
Abstract
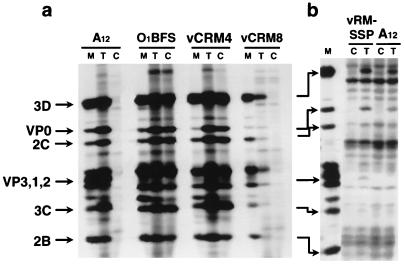
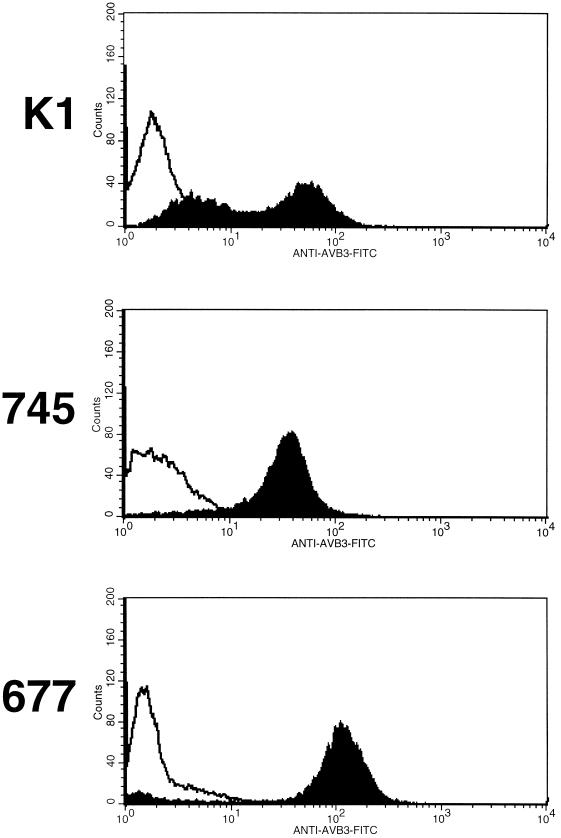
Adsorption and plaque formation of foot-and-mouth disease virus (FMDV) serotype A12 are inhibited by antibodies to the integrin alpha(v)beta3 (A. Berinstein et al., J. Virol. 69:2664-2666, 1995). A human cell line, K562, which does not normally express alpha(v)beta3 cannot replicate this serotype unless cells are transfected with cDNAs encoding this integrin (K562-alpha(v)beta3 cells). In contrast, we found that a tissue culture-propagated FMDV, type O1BFS, was able to replicate in nontransfected K562 cells, and replication was not inhibited by antibodies to the endogenously expressed integrin alpha5beta1. A recent report indicating that cell surface heparan sulfate (HS) was required for efficient infection of type O1 (T. Jackson et al., J. Virol. 70:5282-5287, 1996) led us to examine the role of HS and alpha(v)beta3 in FMDV infection. We transfected normal CHO cells, which express HS but not alpha(v)beta3, and two HS-deficient CHO cell lines with cDNAs encoding human alpha(v)beta3, producing a panel of cells that expressed one or both receptors. In these cells, type A12 replication was dependent on expression of alpha(v)beta3, whereas type O1BFS replicated to high titer in normal CHO cells but could not replicate in HS-deficient cells even when they expressed alpha(v)beta3. We have also analyzed two genetically engineered variants of type O1Campos, vCRM4, which has greatly reduced virulence in cattle and can bind to heparin-Sepharose columns, and vCRM8, which is highly virulent in cattle and cannot bind to heparin-Sepharose. vCRM4 replicated in wild-type K562 cells and normal, nontransfected CHO (HS+ alpha(v)beta3-) cells, whereas vCRM8 replicated only in K562 and CHO cells transfected with alpha(v)beta3 cDNAs. A similar result was also obtained in assays using a vCRM4 virus with an engineered RGD-->KGE mutation. These results indicate that virulent FMDV utilizes the alpha(v)beta3 integrin as a primary receptor for infection and that adaptation of type O1 virus to cell culture results in the ability of the virus to utilize HS as a receptor and a concomitant loss of virulence.
Figures

Similar articles
-
Cell recognition by foot-and-mouth disease virus that lacks the RGD integrin-binding motif: flexibility in aphthovirus receptor usage.J Virol. 2000 Feb;74(4):1641-7. doi: 10.1128/jvi.74.4.1641-1647.2000. J Virol. 2000. PMID: 10644333 Free PMC article.
-
High-efficiency utilization of the bovine integrin alpha(v)beta(3) as a receptor for foot-and-mouth disease virus is dependent on the bovine beta(3) subunit.J Virol. 2000 Aug;74(16):7298-306. doi: 10.1128/jvi.74.16.7298-7306.2000. J Virol. 2000. PMID: 10906183 Free PMC article.
-
Arginine-glycine-aspartic acid-specific binding by foot-and-mouth disease viruses to the purified integrin alpha(v)beta3 in vitro.J Virol. 1997 Nov;71(11):8357-61. doi: 10.1128/JVI.71.11.8357-8361.1997. J Virol. 1997. PMID: 9343190 Free PMC article.
-
Efficient infection of cells in culture by type O foot-and-mouth disease virus requires binding to cell surface heparan sulfate.J Virol. 1996 Aug;70(8):5282-7. doi: 10.1128/JVI.70.8.5282-5287.1996. J Virol. 1996. PMID: 8764038 Free PMC article.
-
Effects of two amino acid substitutions in the capsid proteins on the interaction of two cell-adapted PanAsia-1 strains of foot-and-mouth disease virus serotype O with heparan sulfate receptor.Virol J. 2014 Jul 24;11:132. doi: 10.1186/1743-422X-11-132. Virol J. 2014. PMID: 25056022 Free PMC article.
Cited by
-
Single Amino Acid Substitutions Surrounding the Icosahedral Fivefold Symmetry Axis Are Critical for Alternative Receptor Usage of Foot-and-Mouth Disease Virus.Viruses. 2020 Oct 9;12(10):1147. doi: 10.3390/v12101147. Viruses. 2020. PMID: 33050303 Free PMC article.
-
Initial evidence on differences among Enterovirus 71, Coxsackievirus A16 and Coxsackievirus B4 in binding to cell surface heparan sulphate.Virusdisease. 2014;25(3):277-84. doi: 10.1007/s13337-013-0172-x. Epub 2013 Dec 4. Virusdisease. 2014. PMID: 25674594 Free PMC article.
-
Evaluation of genetically engineered derivatives of a Chinese strain of foot-and-mouth disease virus reveals a novel cell-binding site which functions in cell culture and in animals.J Virol. 2003 Mar;77(5):3269-80. doi: 10.1128/jvi.77.5.3269-3280.2003. J Virol. 2003. PMID: 12584350 Free PMC article.
-
Basic patches on the E2 glycoprotein of eastern equine encephalitis virus influence viral vascular clearance and dissemination in mice.J Virol. 2025 Jun 17;99(6):e0060225. doi: 10.1128/jvi.00602-25. Epub 2025 May 19. J Virol. 2025. PMID: 40387358 Free PMC article.
-
Cell culture propagation of foot-and-mouth disease virus: adaptive amino acid substitutions in structural proteins and their functional implications.Virus Genes. 2020 Feb;56(1):1-15. doi: 10.1007/s11262-019-01714-7. Epub 2019 Nov 27. Virus Genes. 2020. PMID: 31776851 Free PMC article. Review.
References
-
- Acharya R, Fry E, Stuart D, Fox G, Rowlands D, Brown F. The three-dimensional structure of foot-and-mouth disease virus at 2.9 Å resolution. Nature. 1989;337:709–716. - PubMed
-
- Amadori M, Berneri C, Archetti I L. Immunogenicity of foot-and-mouth disease virus grown in BHK-21 suspension cells. Correlation with cell ploidy alterations and abnormal expression of the α5β1 integrin. Vaccine. 1994;12:159–166. - PubMed
-
- Ashida M, Hamada C. Molecular cloning of the hepatitis A virus receptor from a simian cell line. J Gen Virol. 1997;78:1565–1569. - PubMed
-
- Baxt B. Effect of lysosomotropic compounds on early events in foot-and-mouth disease virus replication. Virus Res. 1987;7:257–271. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
