

Monoclonal antibodies to distinct sites on herpes simplex virus (HSV) glycoprotein D block HSV binding to HVEM
- PMID: 9557640
- PMCID: PMC109580
- DOI: 10.1128/JVI.72.5.3595-3601.1998
Monoclonal antibodies to distinct sites on herpes simplex virus (HSV) glycoprotein D block HSV binding to HVEM
Abstract
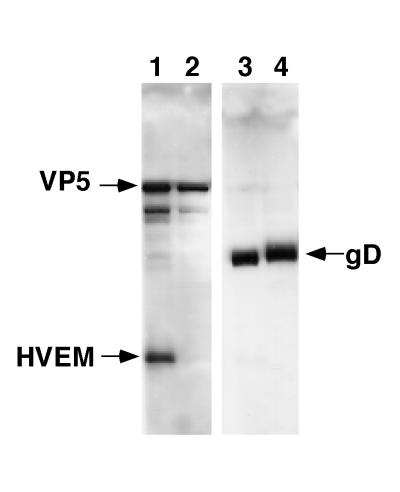
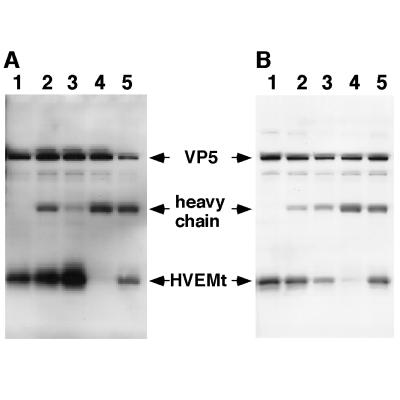
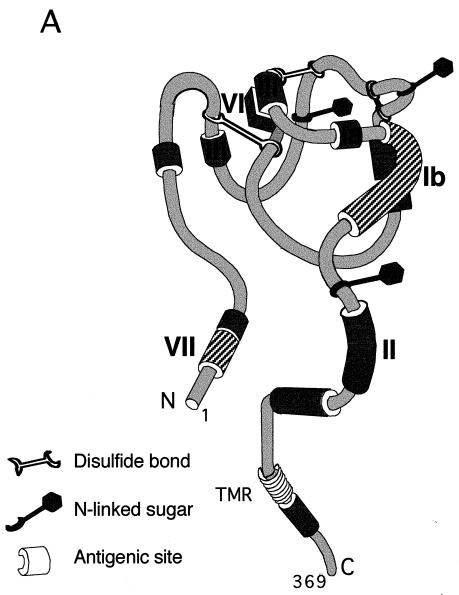
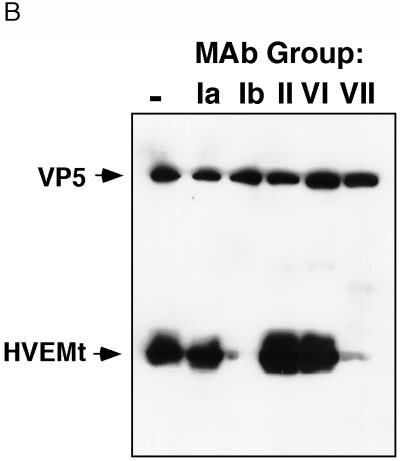
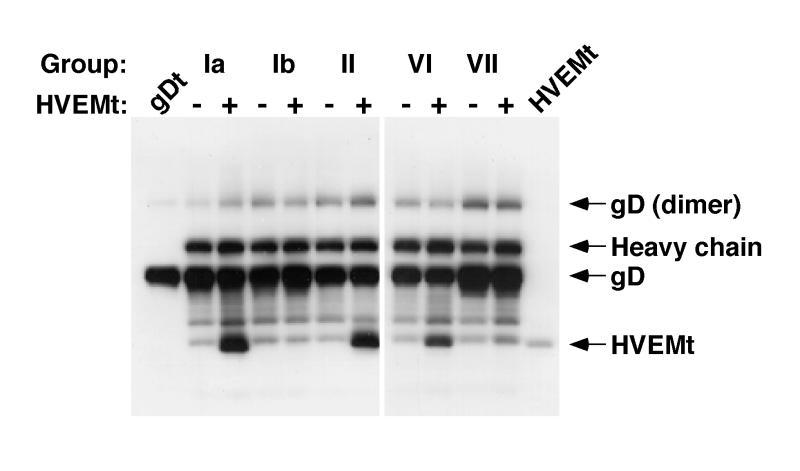
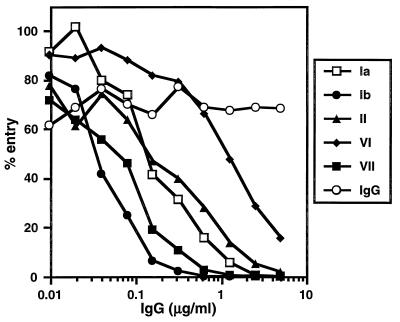
HVEM (for herpesvirus entry mediator) is a member of the tumor necrosis factor receptor superfamily and mediates entry of many strains of herpes simplex virus (HSV) into normally nonpermissive Chinese hamster ovary (CHO) cells. We used sucrose density centrifugation to demonstrate that purified HSV-1 KOS virions bind directly to a soluble, truncated form of HVEM (HVEMt) in the absence of any other cell-associated components. Therefore, HVEM mediates HSV entry by serving as a receptor for the virus. We previously showed that soluble, truncated forms of HSV glycoprotein D (gDt) bind to HVEMt in vitro. Here we show that antibodies specific for gD, but not the other entry glycoproteins gB, gC, or the gH/gL complex, completely block HSV binding to HVEM. Thus, virion gD is the principal mediator of HSV binding to HVEM. To map sites on virion gD which are necessary for its interaction with HVEM, we preincubated virions with gD-specific monoclonal antibodies (MAbs). MAbs that recognize antigenic sites Ib and VII of gD were the only MAbs which blocked the HSV-HVEM interaction. MAbs from these two groups failed to coprecipitate HVEMt in the presence of soluble gDt, whereas the other anti-gD MAbs coprecipitated HVEMt and gDt. Previous mapping data indicated that site VII includes amino acids 11 to 19 and site Ib includes 222 to 252. The current experiments indicate that these sites contain residues important for HSV binding to HVEM. Group Ib and VII MAbs also blocked HSV entry into HVEM-expressing CHO cells. These results suggest that the mechanism of neutralization by these MAbs is via interference with the interaction between gD in the virus and HVEM on the cell. Group Ia and II MAbs failed to block HSV binding to HVEM yet still neutralized HVEM-mediated entry, suggesting that these MAbs block entry at a step other than HVEM binding.
Figures
References
-
- Baker S J, Reddy E P. Transducers of life and death: TNF receptor superfamily and associated proteins. Oncogene. 1996;12:1–9. - PubMed
-
- Bosch D L, Geerligs H J, Weijer W J, Feijlbrief M, Welling G W, Welling-Wester S. Structural properties and reactivity of N-terminal synthetic peptides of herpes simplex virus type 1 glycoprotein D by using antipeptide antibodies and group VII monoclonal antibodies. J Virol. 1987;61:3607–3611. - PMC - PubMed
-
- Brunetti C R, Burke R L, Kornfeld S, Gregory W, Masiarz F R, Dingwell K S, Johnson D C. Herpes simplex virus glycoprotein D acquires mannose 6-phosphate residues and binds to mannose 6-phosphate receptors. J Biol Chem. 1994;269:17067–17074. - PubMed
-
- Buckmaster E A, Gompels U, Minson A. Characterisation and physical mapping of an HSV-1 glycoprotein of approximately 115 × 10(3) molecular weight. Virology. 1984;139:408–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
