

The role of nucleocapsid and U5 stem/A-rich loop sequences in tRNA(3Lys) genomic placement and initiation of reverse transcription in human immunodeficiency virus type 1
- PMID: 9557676
- PMCID: PMC109616
- DOI: 10.1128/JVI.72.5.3907-3915.1998
The role of nucleocapsid and U5 stem/A-rich loop sequences in tRNA(3Lys) genomic placement and initiation of reverse transcription in human immunodeficiency virus type 1
Abstract
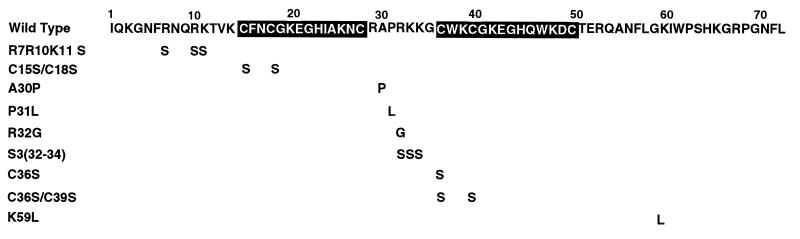
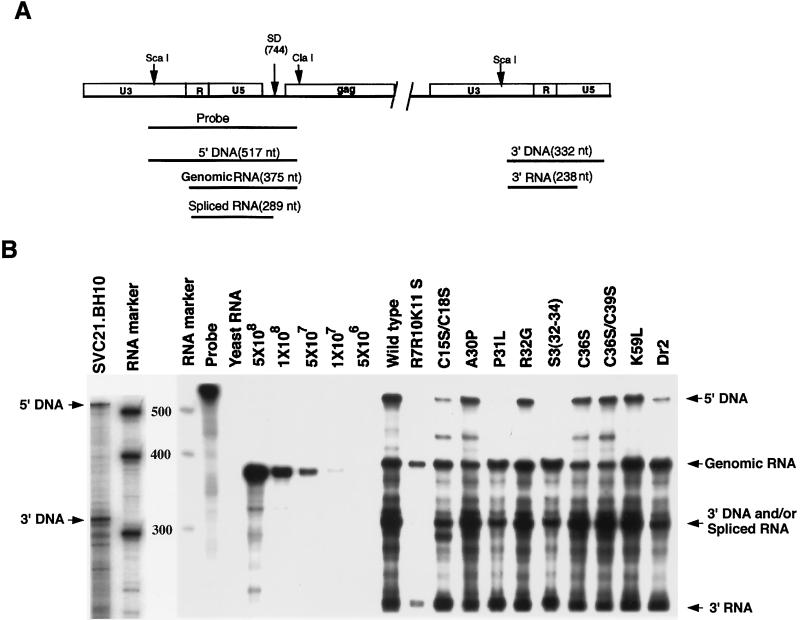
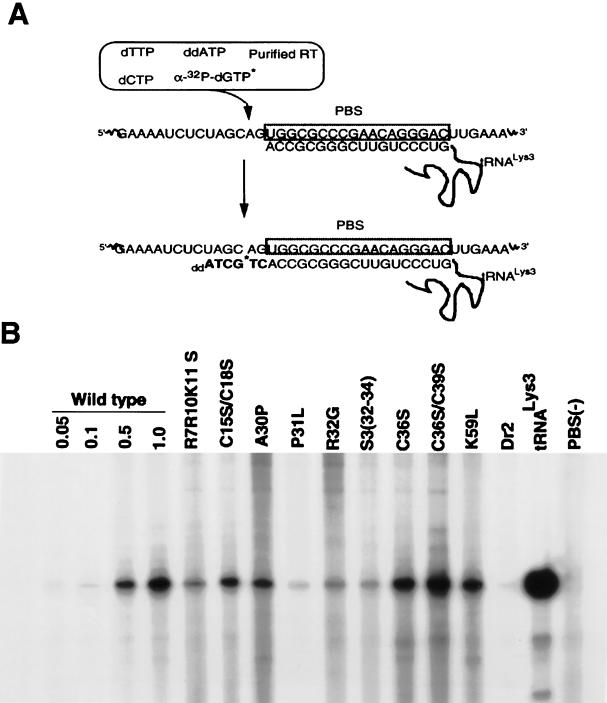
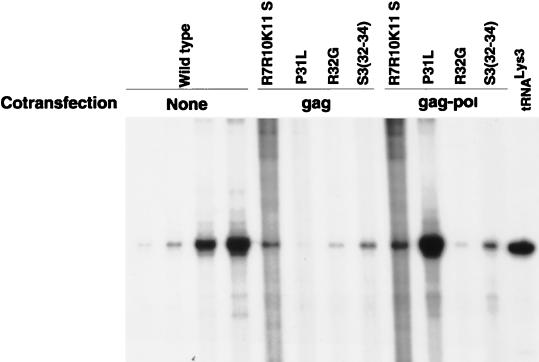
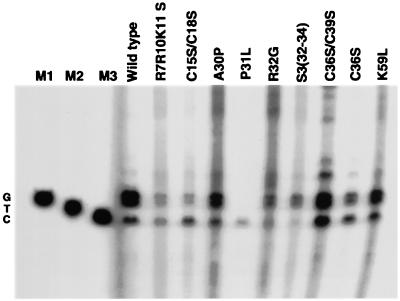
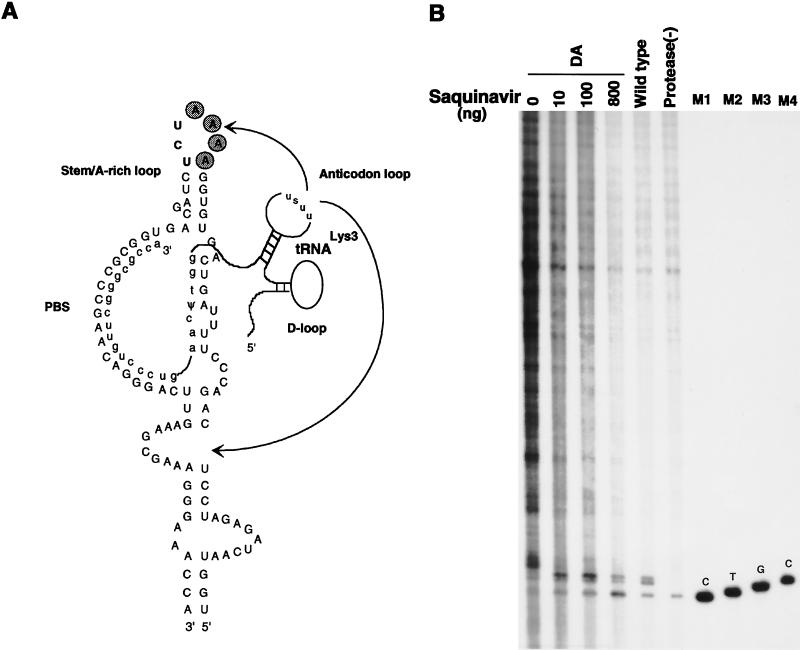
We have studied the effect of mutations in the human immunodeficiency virus type 1 (HIV-1) nucleocapsid (NC) sequence on tRNA(3Lys) genomic placement, i.e., the in vivo placement of primer tRNA(3Lys) on the HIV-1 primer binding site (PBS). HIV-1 produced from COS cells transfected with wild-type or mutant proviral DNA was used in this study. We have found that mutations in the amino acid sequences flanking the first Cys-His box in the NC sequence produce the maximum inhibition of genomic placement. A similar finding was obtained when the NC-facilitated annealing of primer tRNA(3Lys) to the HIV PBS in vitro was studied. However, since the genomic placement of tRNA(3Lys) occurs independently of precursor protein processing, the NC mutations studied here have probably exerted their effect through one or both of the precursor proteins, Pr55gag and/or Pr160(gag-pol). One mutation in the linker region between the two Cys-His boxes, P31L, prevented packaging of both Pr160(gag-pol) and tRNA(3Lys) and prevented the genomic placement of tRNA(3Lys). Both packaging and genomic placement were rescued by cotransfection with a plasmid coding for wild-type Pr160(gag-pol). For other linker mutations [R7R10K11 S, R32G, and S3(32-34)], packaging of Pr160(gag-pol) and tRNA(3Lys) was not affected, but genomic placement was, and placement could not be rescued by cotransfection with plasmids coding for either Pr55gag or Pr160(gag-pol). After placement, the initiation of reverse transcription within extracellular virions is characterized by a 2-base DNA extension of the placed tRNA(3Lys). This process requires precursor processing, and those NC mutations which showed the most inhibition of initiation were in either of the two NC Cys-His boxes. Destabilization of a U5 stem-A-rich loop immediately upstream of the PBS (through deletion of four consecutive A's in the loop) did not affect the in vivo genomic placement of tRNA(3Lys) but resulted in the presence in the extracellular virus of longer cDNA extensions of tRNA(3Lys), with a corresponding decrease in the presence of unextended and 2-base-extended tRNA(3Lys).
Figures
References
-
- Arts E J, Ghosh M, Jacques P S, Ehresmann B, LeGrice S F J. Restoration of tRNALys3-primed (−) strand DNA synthesis to an HIV-1 reverse transcriptase mutant with extended tRNAs. J Biol Chem. 1996;271:9054–9061. - PubMed
-
- Arts E J, Stetor S R, Li X, Rausch J W, Howard K J, Ehresmann B, North T W, Wohrl B M, Goody R S, Wainberg M A, LeGrice S F J. Initiation of (−) strand DNA synthesis from the tRNALys3 on lentiviral RNAs: implications of specific HIV-1 RNA-tRNALys3 interactions inhibiting primer utilization by retroviral reverse transcriptases. Proc Natl Acad Sci USA. 1996;93:10063–10068. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
