

Detection of a novel bovine lymphotropic herpesvirus
- PMID: 9557713
- PMCID: PMC109653
- DOI: 10.1128/JVI.72.5.4237-4242.1998
Detection of a novel bovine lymphotropic herpesvirus
Abstract
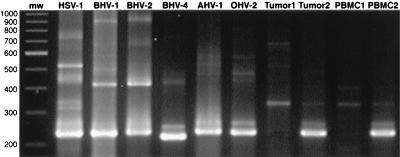
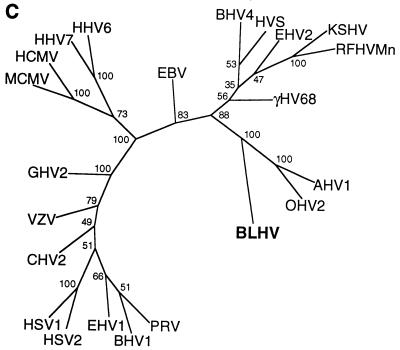
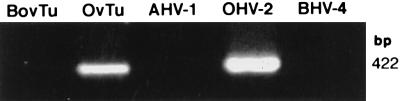
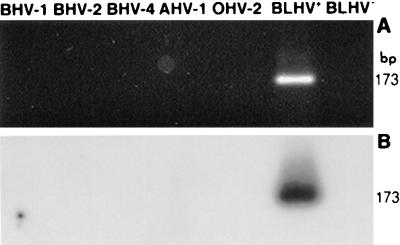
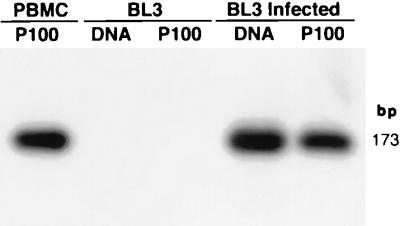
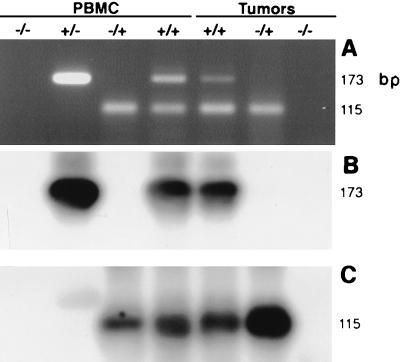
Degenerate PCR primers which amplify a conserved region of the DNA polymerase genes of the herpesvirus family were used to provide sequence evidence for a new bovine herpesvirus in bovine B-lymphoma cells and peripheral blood mononuclear cells (PBMC). The sequence of the resultant amplicon was found to be distinct from those of known herpesvirus isolates. Alignment of amino acid sequences demonstrated 70% identity with ovine herpesvirus 2, 69% with alcelaphine herpesvirus 1, 65% with bovine herpesvirus 4, and 42% with bovine herpesvirus 1. Phylogenetic analysis placed this putative virus within the tumorigenic Gammaherpesvirinae subfamily, and it is tentatively identified as bovine lymphotropic herpesvirus. This novel agent was expressed in vitro from infected PBMC, and cell-free supernatants were used to transfer infection to a bovine B-cell line, BL3. Analysis, with specific PCR primers, of DNA from bovine PBMC and lymphoma cells identified infection in blood of 91% of adult animals (n = 101), 63% of lymphomas (n = 32), and 38% of juveniles (n = 13). Of the adults, herpesvirus infection was present in 94% of animals that were seropositive for bovine leukemia virus (BLV) (n = 63) and in 87% of BLV-seronegative animals (n = 38). Of the seropositive group, 17 animals exhibited persistent lymphocytosis, and 100% of these were herpesvirus positive by PCR. A role for bovine lymphotropic herpesvirus as a cofactor in BLV pathogenesis is considered.
Figures

References
-
- Baxter S I F, Pow I, Bridgen A, Reid H W. PCR detection of the sheep-associated agent of malignant catarrhal fever. Arch Virol. 1993;132:145–159. - PubMed
-
- Cockerell G L, Rovnak J. The correlation between the direct and indirect detection of bovine leukemia virus infection in cattle. Leukocyte Res. 1988;12:465–469. - PubMed
-
- Degen J L, Neubauer M G, Degen S J, Seyfried C E, Morris D R. Regulation of protein synthesis in mitogen-activated bovine lymphocytes. Analysis of actin-specific and total mRNA accumulation and utilization. J Biol Chem. 1983;258:12153–12162. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
