

Regulation of the zinc transporter ZnT-1 by dietary zinc
- PMID: 9560190
- PMCID: PMC20175
- DOI: 10.1073/pnas.95.9.4841
Regulation of the zinc transporter ZnT-1 by dietary zinc
Abstract
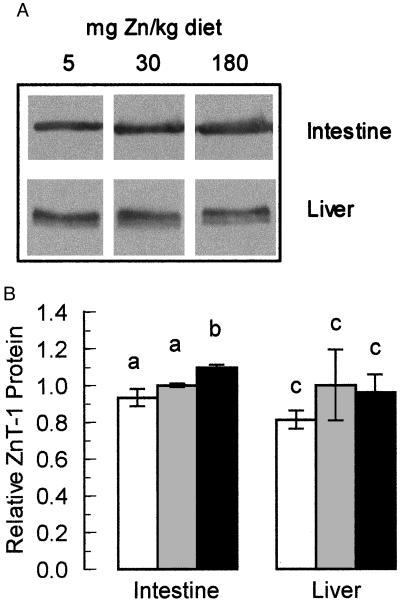
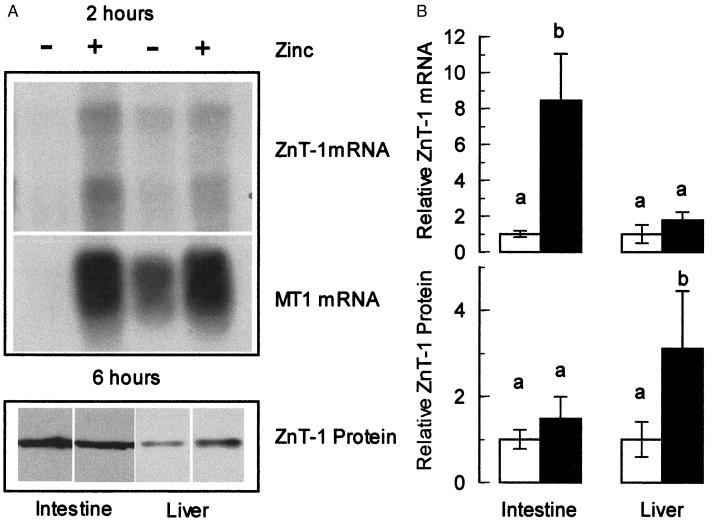
The understanding of mechanisms controlling zinc absorption and metabolism at the molecular level has advanced recently. Kinetics of zinc transport have been investigated for many years, but only recently have genes coding for proteins thought to be involved in the transport process been cloned. Four putative zinc transporters, known as ZnT-1 through ZnT-4, have now been described. Among these transporters, only ZnT-1 is ubiquitously expressed. In this report, we examine the pattern of ZnT-1 expression in the intestine and analyze the regulation of ZnT-1 by dietary zinc in both the intestine and liver. Immunofluorescence demonstrated that intestinal ZnT-1 was most abundant at the basolateral surface of enterocytes lining the villi of the duodenum and jejunum. By Western blot analysis, intestinal and liver ZnT-1 protein migrated as a 42- and 36-kDa protein, respectively. Dietary zinc supplementation elevated the level of intestinal ZnT-1 mRNA and protein approximately 50% and 10%, respectively, but had no effect in the liver. In response to an acute oral zinc dose, the level of intestinal ZnT-1 mRNA was up-regulated 8-fold, without a corresponding increase in ZnT-1 protein. Conversely, the acute oral dose did not affect liver ZnT-1 mRNA, but resulted in a 5-fold increase in liver ZnT-1 protein. These results represent studies on the expression of intestinal and hepatic ZnT-1 in an intact animal model. The data suggest that ZnT-1 is at least part of the mechanism by which dietary zinc is absorbed and that, despite the zinc responsiveness of the ZnT-1 gene, additional factors may be regulating the steady-state level of ZnT-1 transporter protein.
Figures
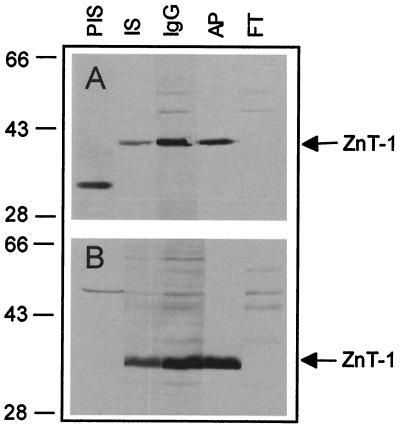
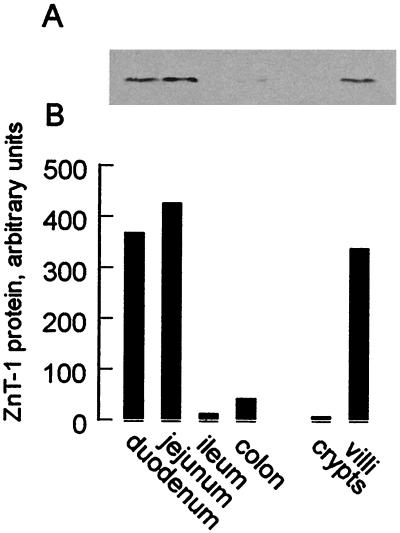
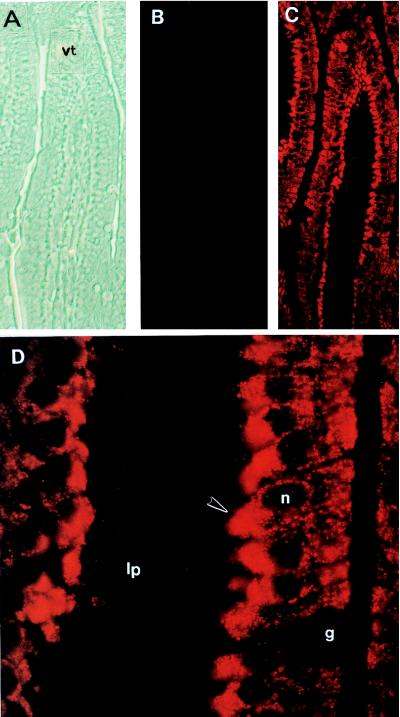
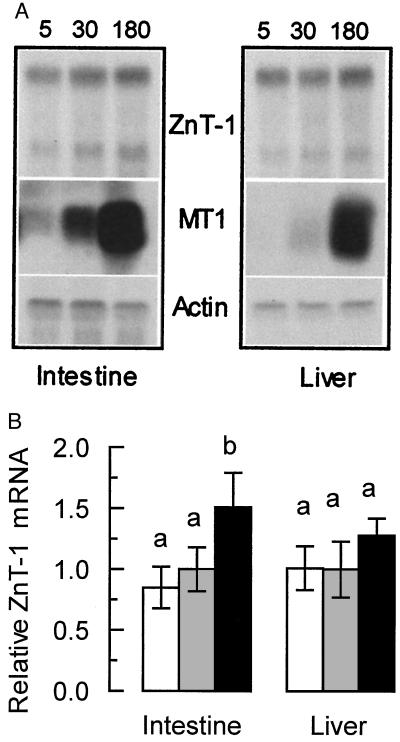
References
-
- Cousins R J. In: Present Knowledge in Nutrition. 7th Ed. Filer L J, Ziegler E E, editors. Washington, DC: Int. Life Sci. Inst. Nutr. Foundation; 1996. pp. 293–306.
-
- Steel L, Cousins R J. Am J Physiol. 1985;248:G46–G53. - PubMed
-
- Hoadley J E, Leinart A S, Cousins R J. Am J Physiol. 1987;252:G825–G831. - PubMed
-
- Raffaniello R D, Lee S Y, Teichberg S, Wapnir R A. J Cell Physiol. 1992;152:356–361. - PubMed
-
- Ackland M L, Danks D M, McArdle H J. J Cell Physiol. 1988;135:521–526. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
