

Crystal structures of two tropinone reductases: different reaction stereospecificities in the same protein fold
- PMID: 9560196
- PMCID: PMC20181
- DOI: 10.1073/pnas.95.9.4876
Crystal structures of two tropinone reductases: different reaction stereospecificities in the same protein fold
Abstract
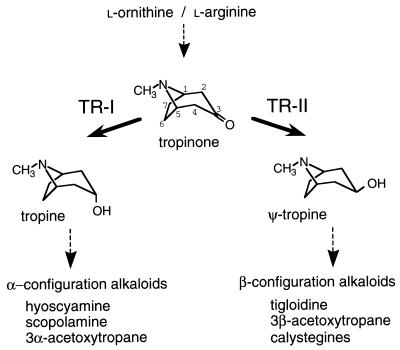
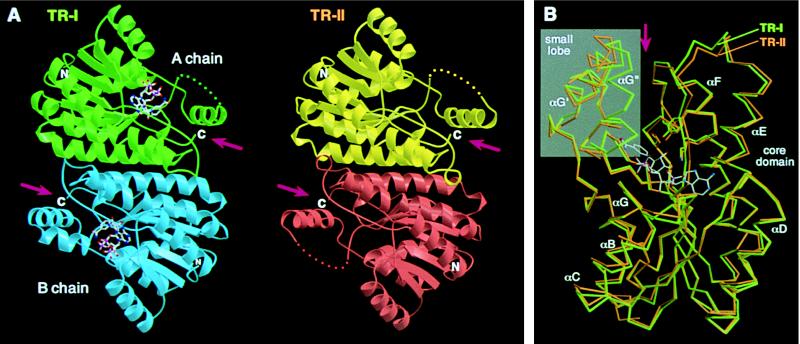
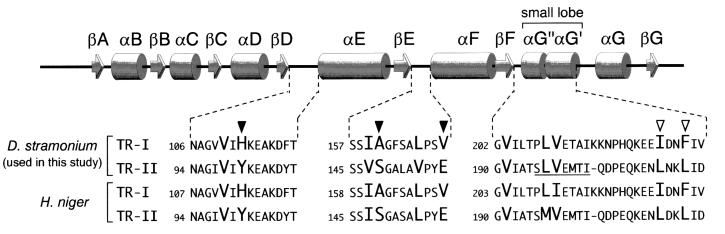
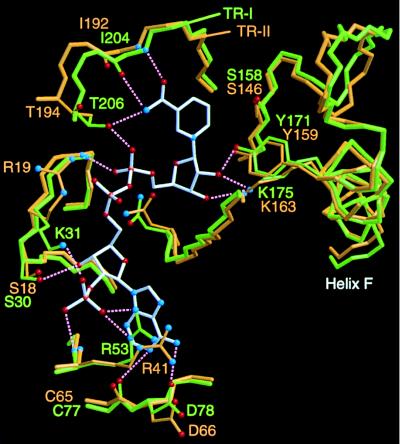
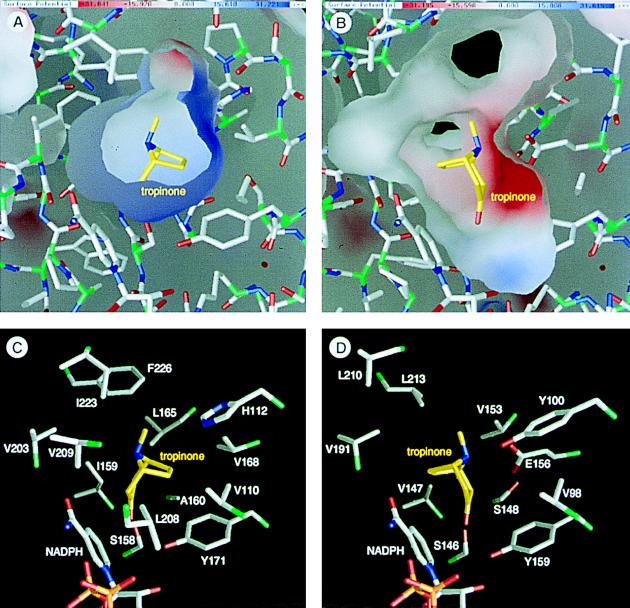
A pair of tropinone reductases (TRs) share 64% of the same amino acid residues and belong to the short-chain dehydrogenase/reductase family. In the synthesis of tropane alkaloids in several medicinal plants, the TRs reduce a carbonyl group of an alkaloid intermediate, tropinone, to hydroxy groups with different diastereomeric configurations. To clarify the structural basis for their different reaction stereospecificities, we determined the crystal structures of the two enzymes at 2.4- and 2.3-A resolutions. The overall folding of the two enzymes was almost identical. The conservation was not confined within the core domains that are conserved within the protein family but extended outside the core domain where each family member has its characteristic structure. The binding sites for the cofactor and the positions of the active site residues were well conserved between the two TRs. The substrate binding site was composed mostly of hydrophobic amino acids in both TRs, but the presence of different charged residues conferred different electrostatic environments on the two enzymes. A modeling study indicated that these charged residues play a major role in controlling the binding orientation of tropinone within the substrate binding site, thereby determining the stereospecificity of the reaction product. The results obtained herein raise the possibility that in certain cases different stereospecificities can be acquired in enzymes by changing a few amino acid residues within substrate binding sites.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
