

Mechanism of platelet inhibition by nitric oxide: in vivo phosphorylation of thromboxane receptor by cyclic GMP-dependent protein kinase
- PMID: 9560198
- PMCID: PMC20183
- DOI: 10.1073/pnas.95.9.4888
Mechanism of platelet inhibition by nitric oxide: in vivo phosphorylation of thromboxane receptor by cyclic GMP-dependent protein kinase
Abstract
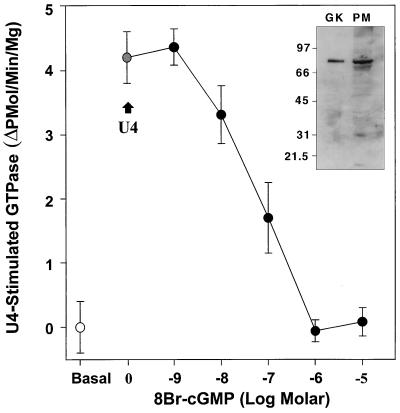
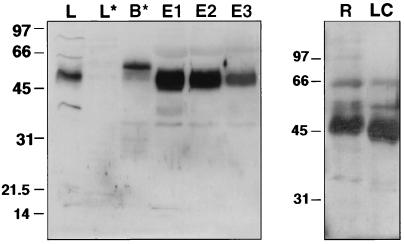
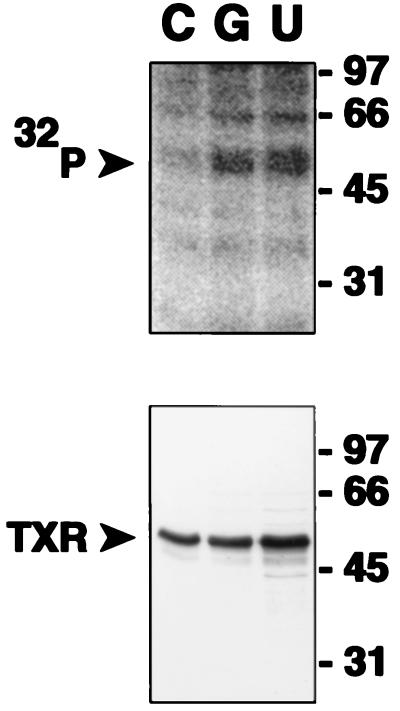
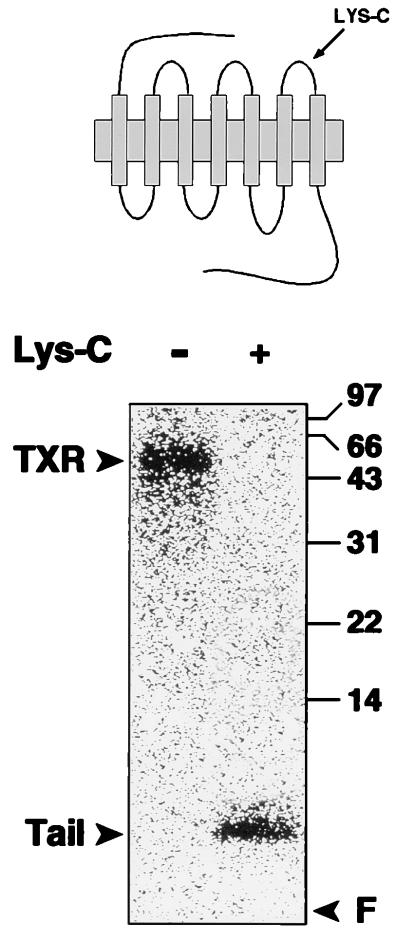
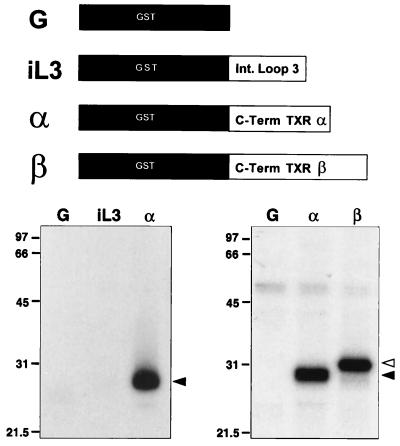
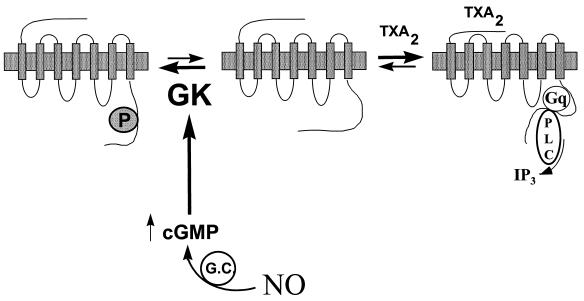
Nitric oxide (NO) is a potent vasodilator and inhibitor of platelet activation. NO stimulates production of cGMP and activates cGMP-dependent protein kinase (G kinase), which by an unknown mechanism leads to inhibition of Galphaq-phospholipase C-inositol 1, 4,5-triphosphate signaling and intracellular calcium mobilization for several important agonists, including thromboxane A2 (TXA2). To explore the mechanism of platelet inhibition by NO, activation of platelet TXA2 receptors in the presence of cGMP was studied. The nonhydrolyzable analog 8-bromo-cyclic GMP (8-Br-cGMP) potently inhibited activation of the TXA2-specific GTPase in platelet membranes in a concentration-dependent fashion, suggesting that G kinase catalyzes the phosphorylation of some proximal component of the receptor-G protein signaling pathway. Nanomolar concentrations of G kinase were found to catalyze the phosphorylation of platelet TXA2 receptors in vitro, but not Galphaq copurifying with the TXA2 receptors in these experiments. Using immunoaffinity methods, in vivo phosphorylation of TXA2 receptors by cyclic GMP was demonstrated from 32P-labeled cells treated with 8-Br-cGMP. Peptide mapping studies of in vivo phosphorylated TXA2 receptors demonstrated cGMP mediates phosphorylation of the carboxyl terminus of the TXA2 receptor. G kinase also catalyzed the phosphorylation of peptides corresponding to the cytoplasmic tails of both alpha and beta forms of the receptor but not control peptide or a peptide corresponding to the third intracytoplasmic loop of the TXA2 receptor. These data identify TXA2 receptors as cGMP-dependent protein kinase substrates and support a novel mechanism for the inhibition of cell function by NO in which activation of G kinase inhibits signaling by G protein-coupled receptors by catalyzing their phosphorylation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
