

The Gfi-1B proto-oncoprotein represses p21WAF1 and inhibits myeloid cell differentiation
- PMID: 9566867
- PMCID: PMC110626
- DOI: 10.1128/MCB.18.5.2462
The Gfi-1B proto-oncoprotein represses p21WAF1 and inhibits myeloid cell differentiation
Abstract
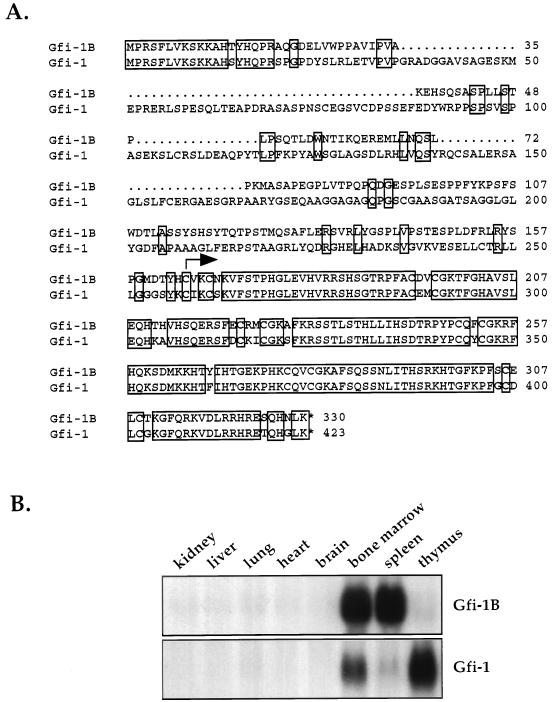
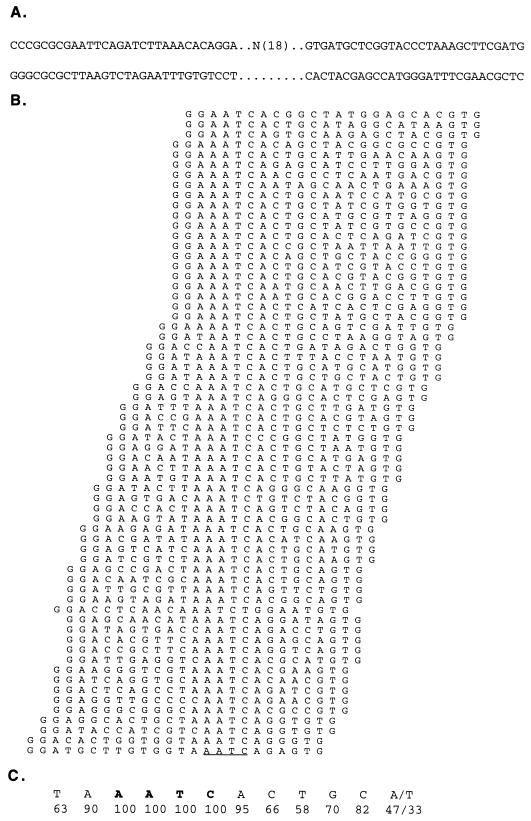
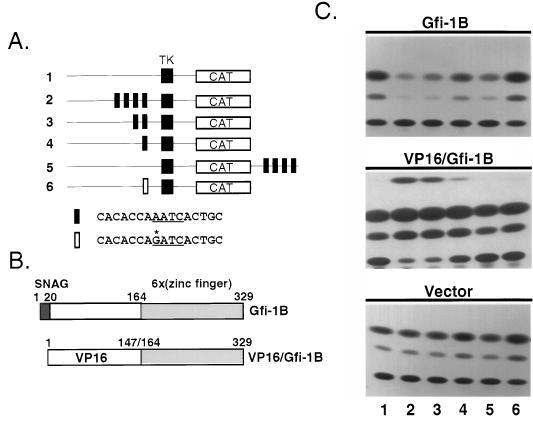
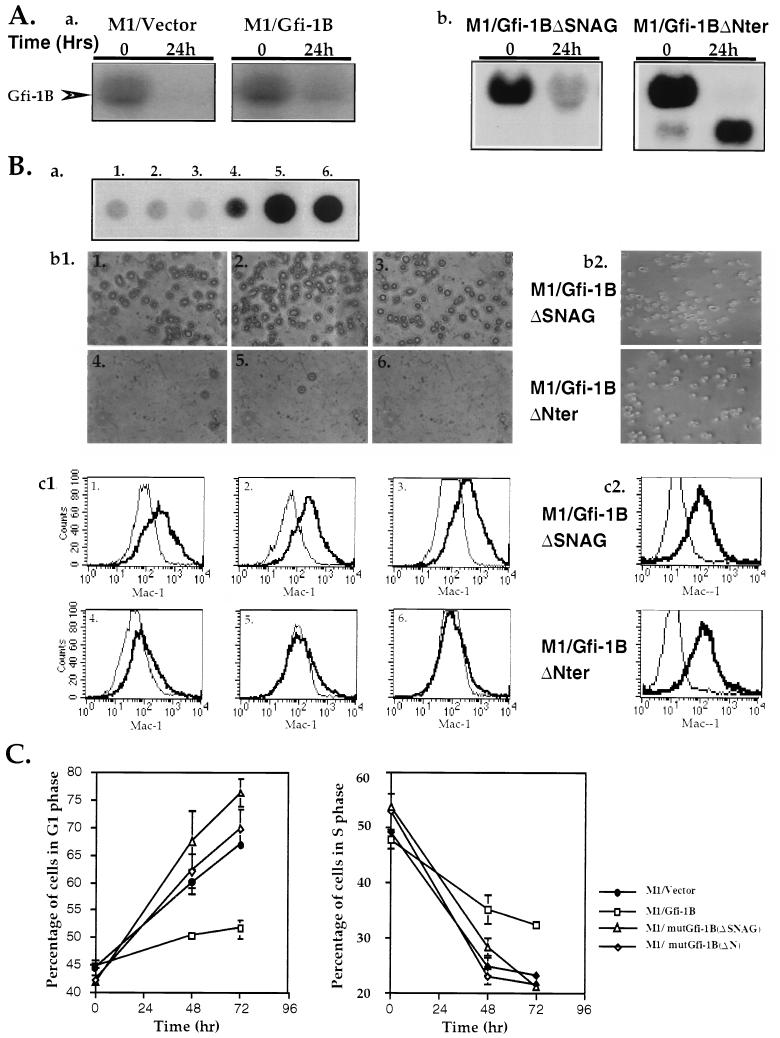
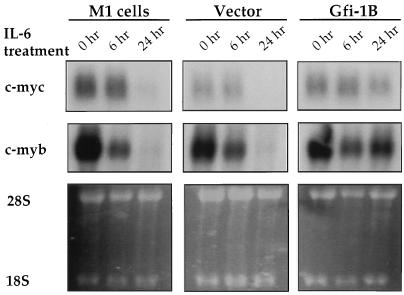
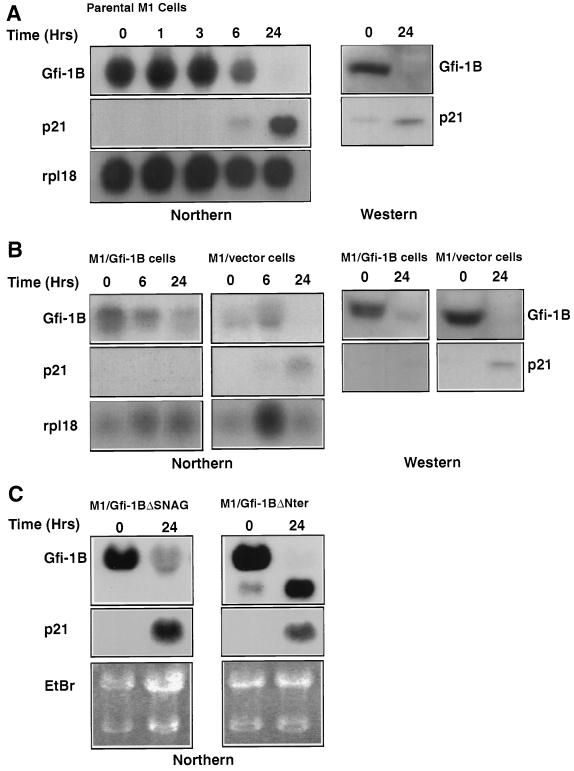
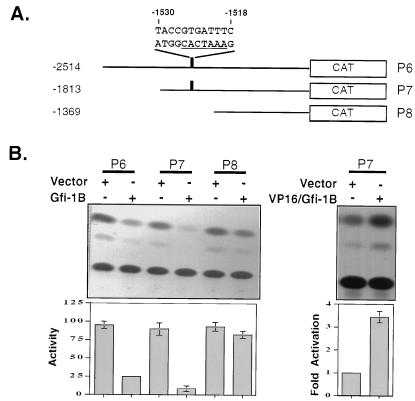
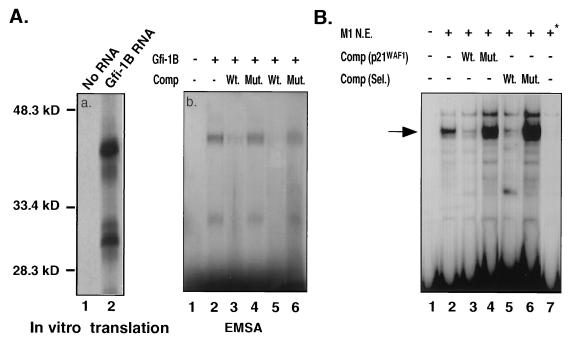
Gfi-1 is a cellular proto-oncogene that was identified as a target of provirus integration in T-cell lymphoma lines selected for interleukin-2 (IL-2) independence in culture and in primary retrovirus-induced lymphomas. Gfi-1 encodes a zinc finger protein that functions as a transcriptional repressor. Here we show that Gfi-1B, a Gfi-1 related gene expressed in bone marrow and spleen, also encodes a transcriptional repressor. IL-6-induced G1 arrest and differentiation of the myelomonocytic cell line M1 were linked to the downregulation of Gfi-1B and the parallel induction of the cyclin-dependent kinase inhibitor p21WAF1. Experiments addressing the potential mechanism of the apparent coordinate regulation of these genes revealed that Gfi-1B represses p21WAF1 directly by binding to a high-affinity site at -1518 to -1530 in the p21WAF1 promoter. Forced expression of Gfi-1B, but not of Gfi-1B deletion mutants lacking the repressor domain, blocked the IL-6-mediated induction of p21WAF1 and inhibited G1 arrest and differentiation. We conclude that Gfi-1B is a direct repressor of the p21WAF1 promoter, the first such repressor identified to date, and that sustained expression of Gfi-1B blocks IL-6-induced G1 arrest and differentiation of M1 cells perhaps because it prevents p21WAF1 induction by IL-6.
Figures
References
-
- Anderson S, Davis D H, Dahlbäck H, Jörnvall H, Russell D W. Cloning, structure and expression of the mitochondrial cytochrome P-450 sterol 26-hydroxylase, a bile biosynthetic enzyme. J Biol Chem. 1989;264:8222–8229. - PubMed
-
- Arnaout M A. Structure and function of the leukocyte adhesion molecules CD11/CD18. Blood. 1990;75:1037–1050. - PubMed
-
- Bell D W, Taguchi T, Jenkins N A, Gilbert D J, Copeland N G, Gilks C B, Zweidler-McKay P, Grimes H L, Tsichlis P N, Testa J R. Chromosomal localization of a gene, GFI1, encoding a novel zinc finger protein reveals a new syntenic region between man and rodents. Cytogenet Cell Genet. 1995;70:263–267. - PubMed
-
- Bies J, Hoffman B, Amanullah A, Giese T, Wolff L. B-Myb prevents growth arrest associated with terminal differentiation of monocytic cells. Oncogene. 1996;12:355–363. - PubMed
-
- Biggs J R, Kudlow J E, Kraft A S. The role of the transcription factor Sp1 in regulating the expression of the WAF1/CIP1 gene in U937 leukemic cells. J Biol Chem. 1996;271:901–906. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases