

Posttranslational regulation of Ty1 retrotransposition by mitogen-activated protein kinase Fus3
- PMID: 9566871
- PMCID: PMC110630
- DOI: 10.1128/MCB.18.5.2502
Posttranslational regulation of Ty1 retrotransposition by mitogen-activated protein kinase Fus3
Erratum in
- Mol Cell Biol 1998 Sep;18(9):5620
Abstract
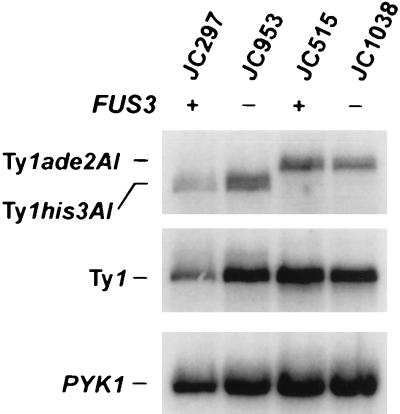
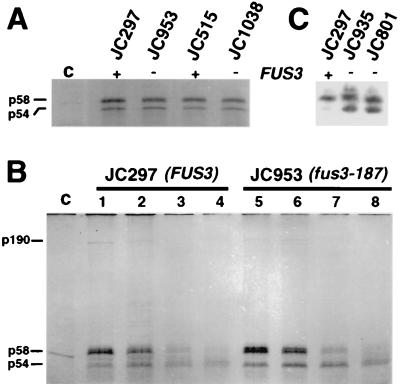
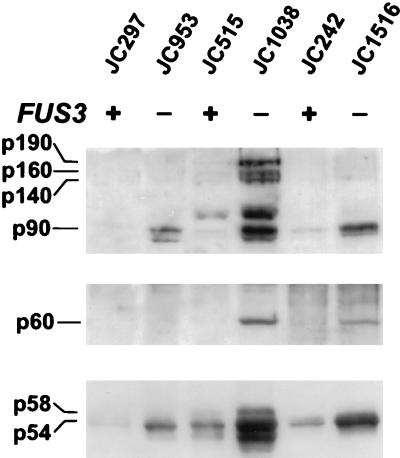
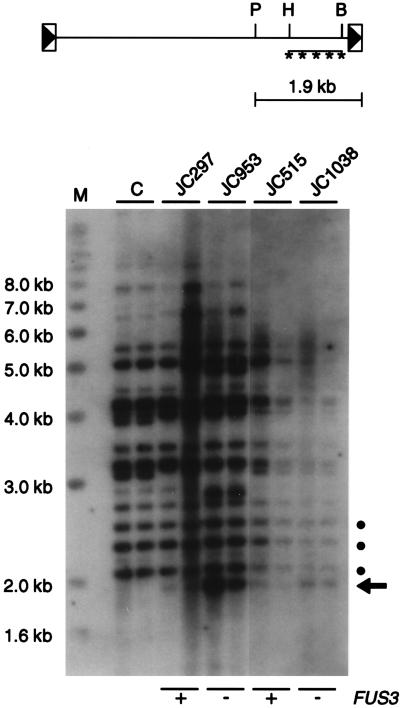
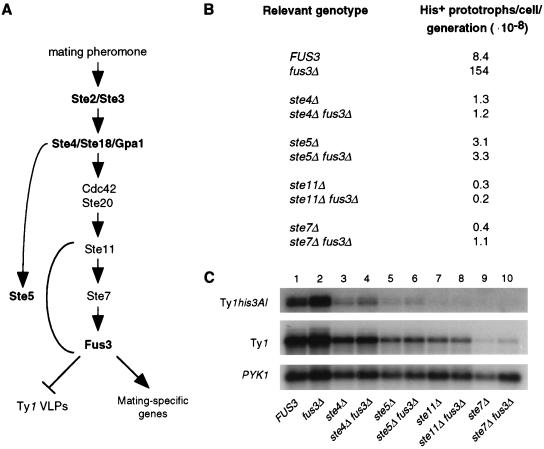
Ty1 retrotransposons in Saccharomyces cerevisiae are maintained in a state of transpositional dormancy. We isolated a mutation, rtt100-1, that increases the transposition of genomic Ty1 elements 18- to 56-fold but has little effect on the transposition of related Ty2 elements. rtt100-1 was shown to be a null allele of the FUS3 gene, which encodes a haploid-specific mitogen-activated protein kinase. In fus3 mutants, the levels of Ty1 RNA, protein synthesis, and proteolytic processing were not altered relative to those in FUS3 strains but steady-state levels of TyA, integrase, and reverse transcriptase proteins and Ty1 cDNA were all increased. These findings suggest that Fus3 suppresses Ty1 transposition by destabilizing viruslike particle-associated proteins. The Fus3 kinase is activated through the mating-pheromone response pathway by phosphorylation at basal levels in naive cells and at enhanced levels in pheromone-treated cells. We demonstrate that suppression of Ty1 transposition in naive cells requires basal levels of Fus3 activation. Substitution of conserved amino acids required for activation of Fus3 derepressed Ty1 transposition. Moreover, epistasis analyses revealed that components of the pheromone response pathway that act upstream of Fus3, including Ste4, Ste5, Ste7, and Ste11, are required for the posttranslational suppression of Ty1 transposition by Fus3. The regulation of Ty1 transposition by Fus3 provides a haploid-specific mechanism through which environmental signals can modulate the levels of retrotransposition.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases