

c-Maf interacts with c-Myb to regulate transcription of an early myeloid gene during differentiation
- PMID: 9566892
- PMCID: PMC110652
- DOI: 10.1128/MCB.18.5.2729
c-Maf interacts with c-Myb to regulate transcription of an early myeloid gene during differentiation
Abstract
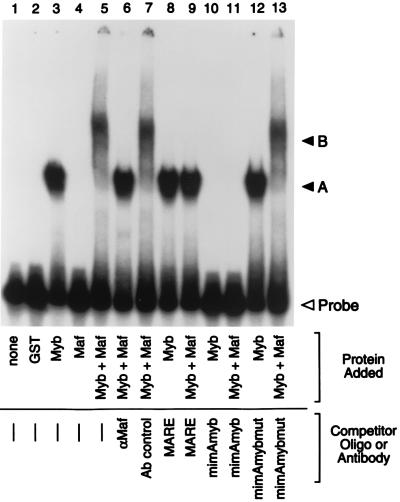
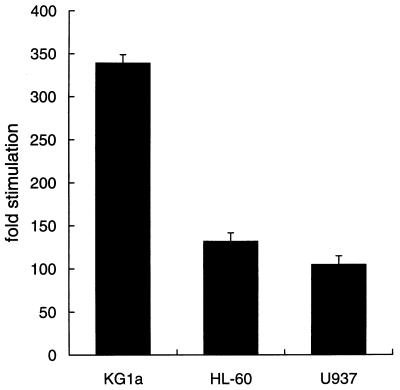
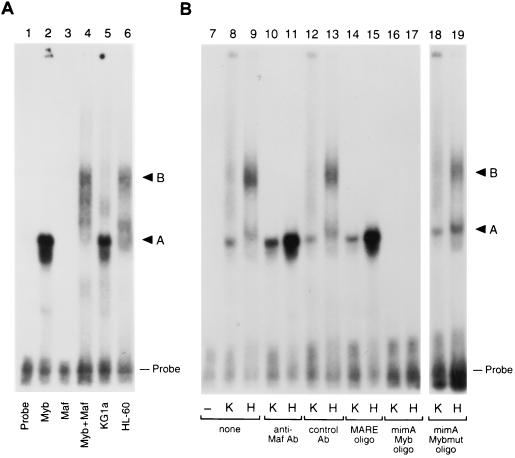
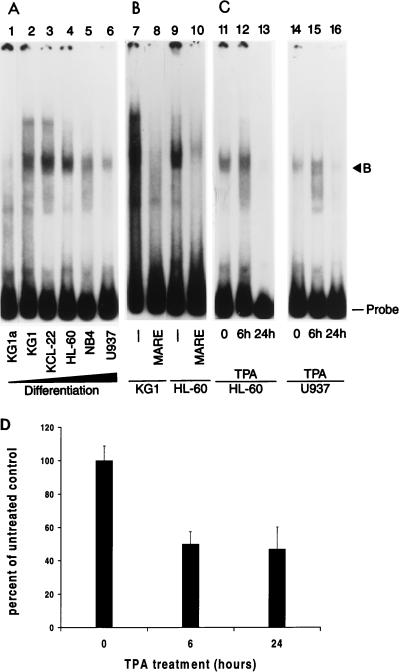
The MafB transcriptional activator plays a pivotal role in regulating lineage-specific gene expression during hematopoiesis by repressing Ets-1-mediated transcription of key erythroid-specific genes in myeloid cells. To determine the effects of Maf family proteins on the transactivation of myeloid-specific genes in myeloid cells, we tested the ability of c-Maf to influence Ets-1- and c-Myb-dependent CD13/APN transcription. Expression of c-Maf in human immature myeloblastic cells inhibited CD13/APN-driven reporter gene activity (85 to 95% reduction) and required the binding of both c-Myb and Ets, but not Maf, to the promoter fragment. c-Maf's inhibition of CD13/APN expression correlates with its ability to physically associate with c-Myb. While c-Maf mRNA and protein levels remain constant during myeloid differentiation, formation of inhibitory Myb-Maf complexes was developmentally regulated, with their levels being highest in immature myeloid cell lines and markedly decreased in cell lines representing later developmental stages. This pattern matched that of CD13/APN reporter gene expression, indicating that Maf modulation of c-Myb activity may be an important mechanism for the control of gene transcription during hematopoietic cell development.
Figures

References
-
- Brelvi Z S, Studzinski G. Coordinate expression of c-myc, c-myb, and histone H4 genes in reversibly differentiating HL-60 cells. J Cell Physiol. 1987;131:43–49. - PubMed
-
- Cordes S P, Barsh G S. The mouse segmentation gene kr encodes a novel basic domain-leucine zipper transcription factor. Cell. 1994;79:1025–1034. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous