

Two distinct types of repression domain in engrailed: one interacts with the groucho corepressor and is preferentially active on integrated target genes
- PMID: 9566899
- PMCID: PMC110659
- DOI: 10.1128/MCB.18.5.2804
Two distinct types of repression domain in engrailed: one interacts with the groucho corepressor and is preferentially active on integrated target genes
Abstract
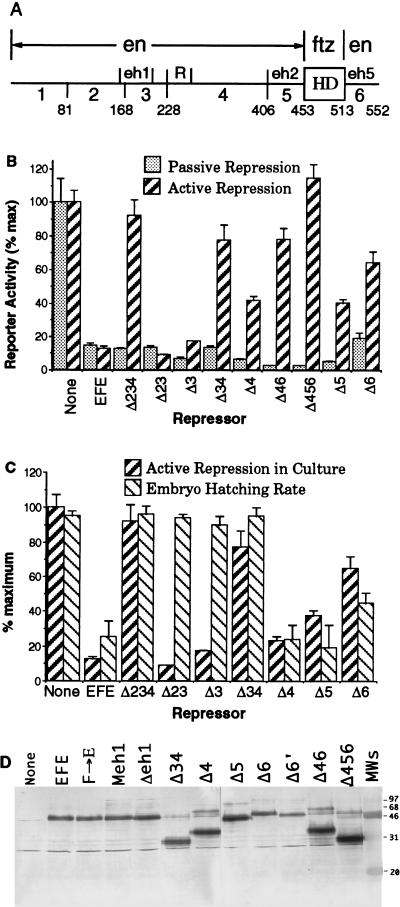
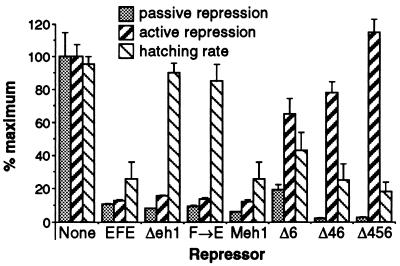
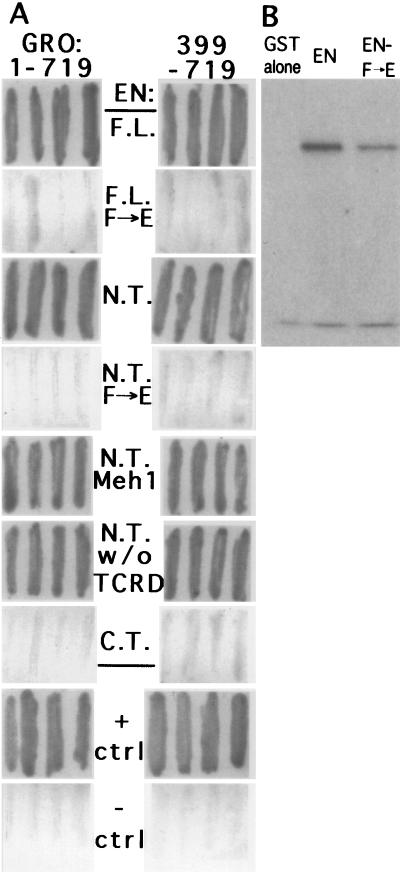
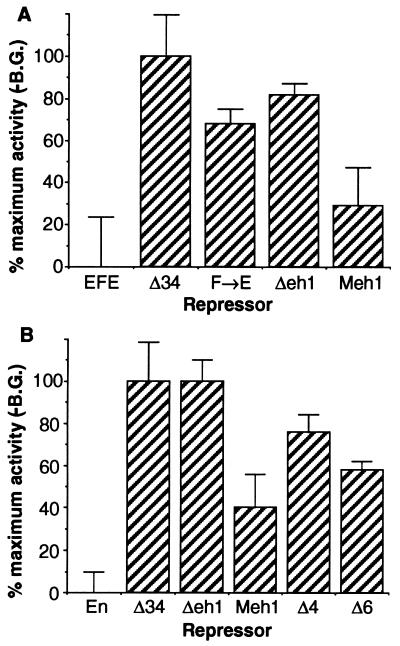
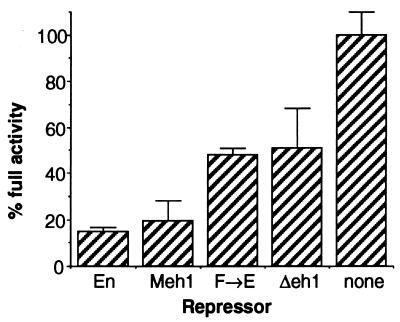
Active transcriptional repression has been characterized as a function of many regulatory factors. It facilitates combinatorial regulation of gene expression by allowing repressors to be dominant over activators under certain conditions. Here, we show that the Engrailed protein uses two distinct mechanisms to repress transcription. One activity is predominant under normal transient transfection assay conditions in cultured cells. A second activity is predominant in an in vivo active repression assay. The domain mediating the in vivo activity (eh1) is highly conserved throughout several classes of homeoproteins and interacts specifically with the Groucho corepressor. While eh1 shows only weak activity in transient transfections, much stronger activity is seen in culture when an integrated target gene is used. In this assay, the relative activities of different repression domains closely parallel those seen in vivo, with eh1 showing the predominant activity. Reducing the amounts of repressor and target gene in a transient transfection assay also increases the sensitivity of the assay to the Groucho interaction domain, albeit to a lesser extent. This suggests that it utilizes rate-limiting components that are relatively low in abundance. Since Groucho itself is abundant in these cells, the results suggest that a limiting component is recruited effectively by the repressor-corepressor complex only on integrated target genes.
Figures

References
-
- Biggin M D, Tjian R. A purified Drosophila homeodomain protein represses transcription in vitro. Cell. 1989;58:433–440. - PubMed
-
- Dorsett D, Viglianti G A, Rutledge B J, Meselson M. Alteration of hsp82 gene expression by the gypsy transposon and suppressor genes in Drosophila melanogaster. Genes Dev. 1989;3:454–468. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases