

Anaphase A chromosome movement and poleward spindle microtubule flux occur At similar rates in Xenopus extract spindles
- PMID: 9566970
- PMCID: PMC2132746
- DOI: 10.1083/jcb.141.3.703
Anaphase A chromosome movement and poleward spindle microtubule flux occur At similar rates in Xenopus extract spindles
Abstract
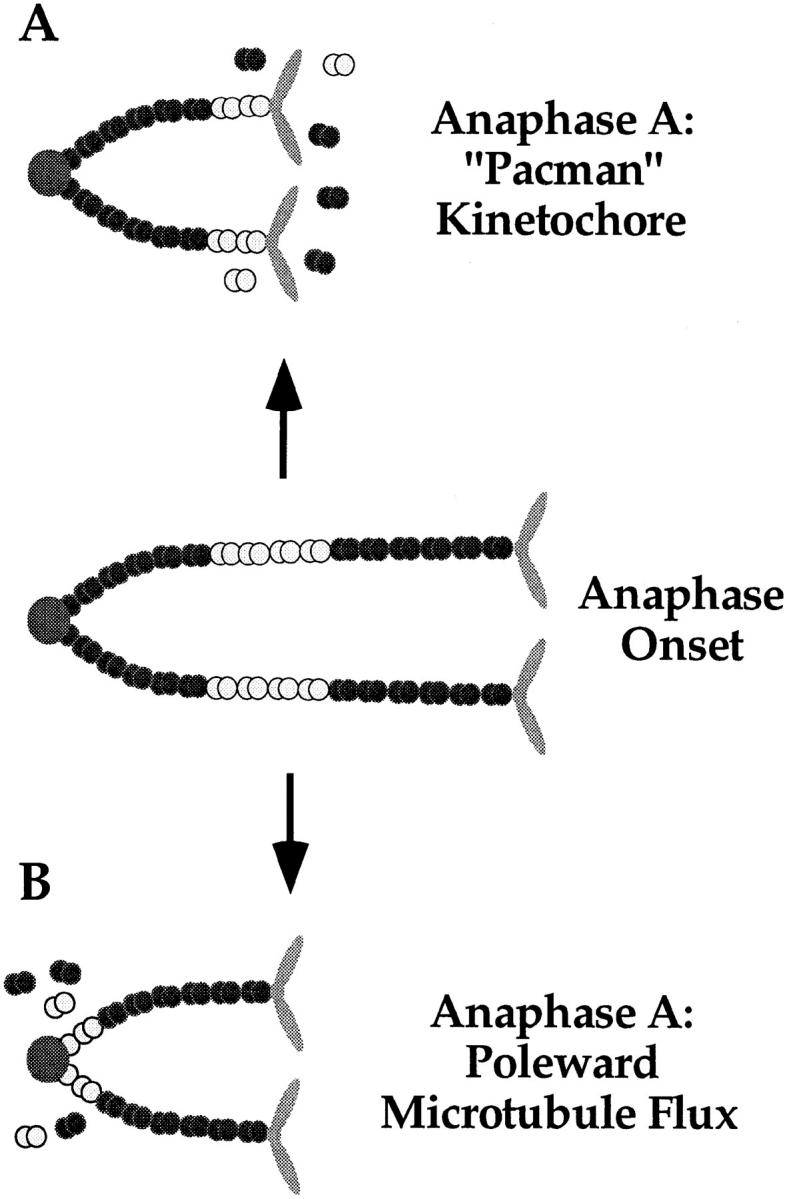
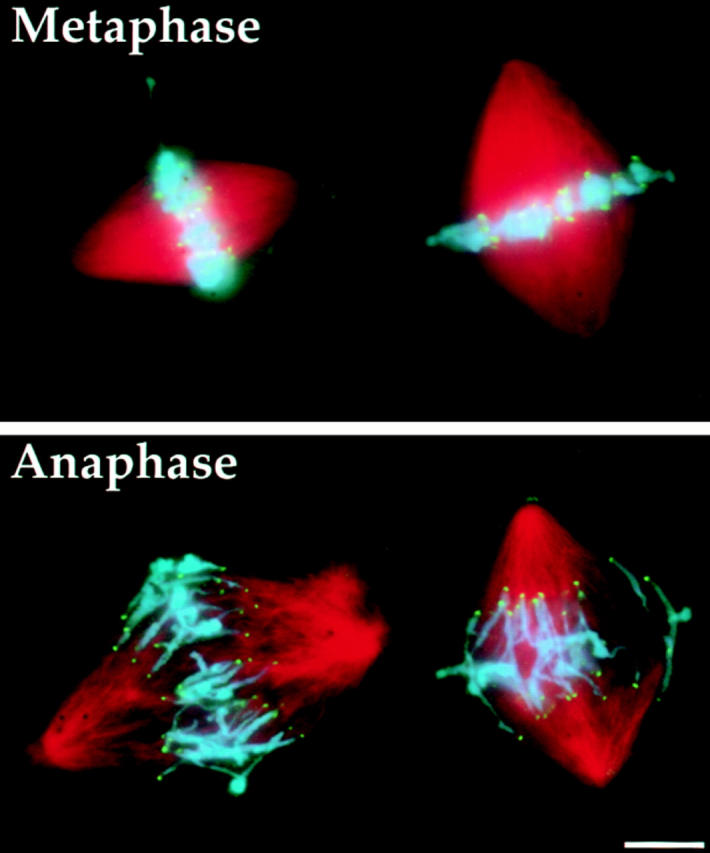
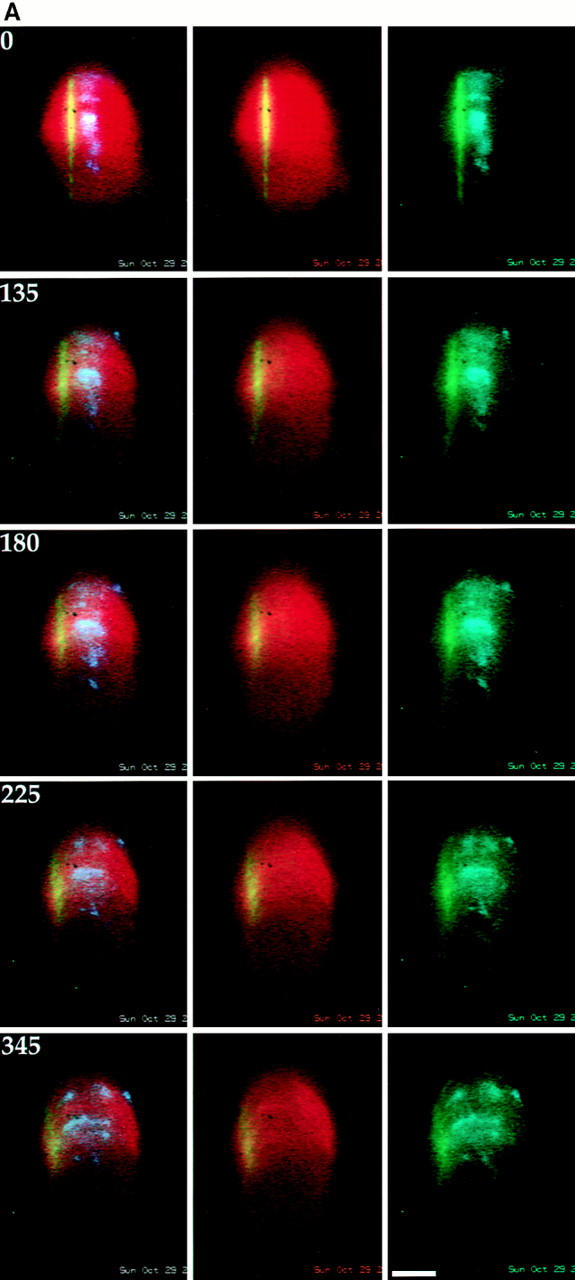
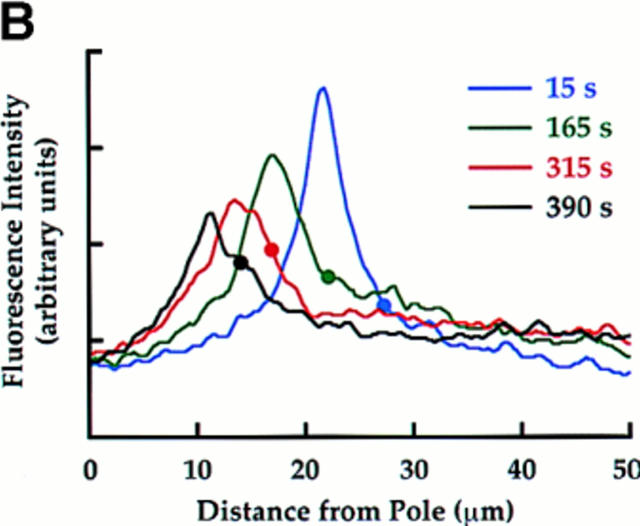
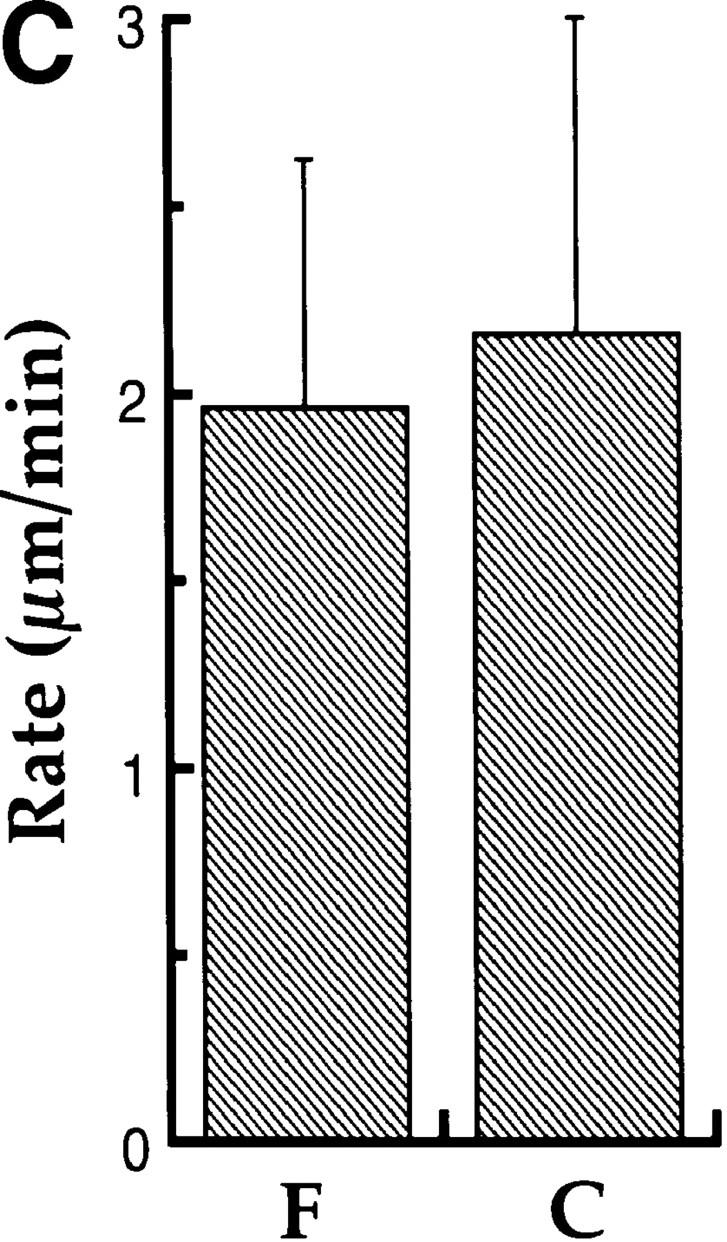
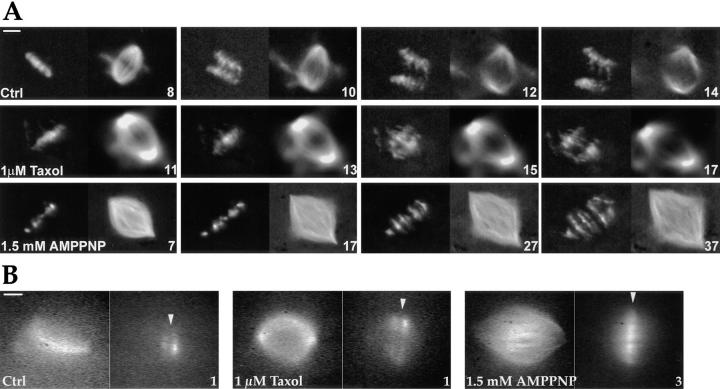
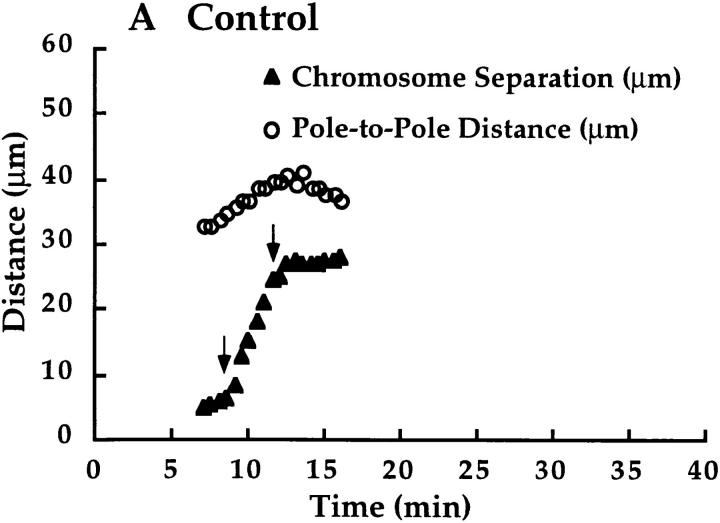
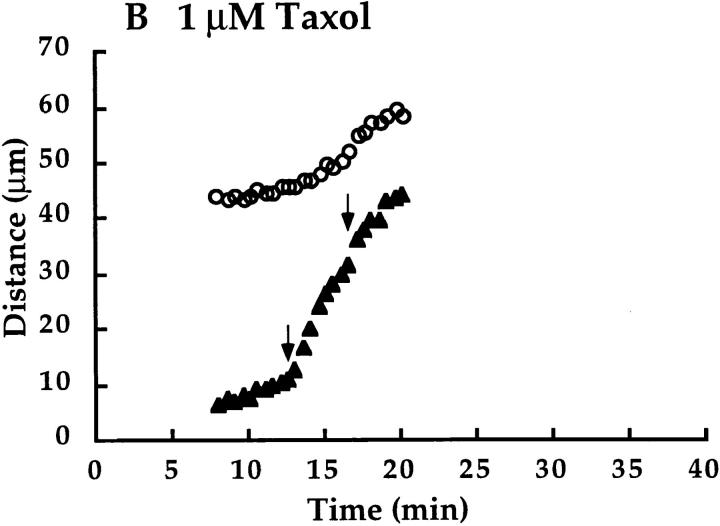
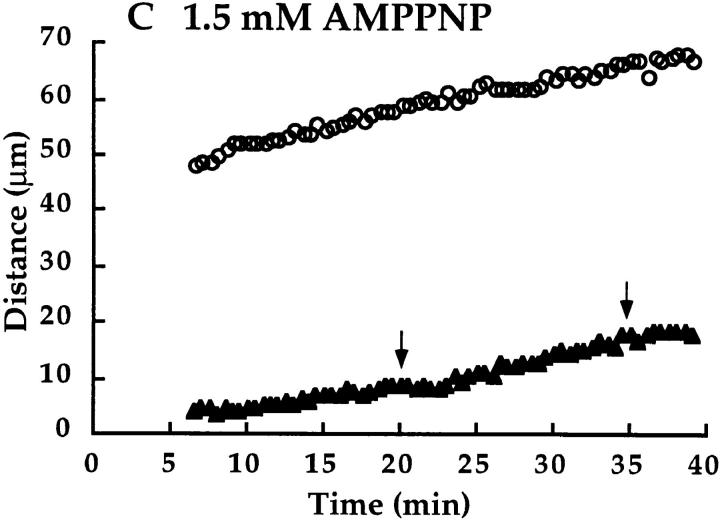
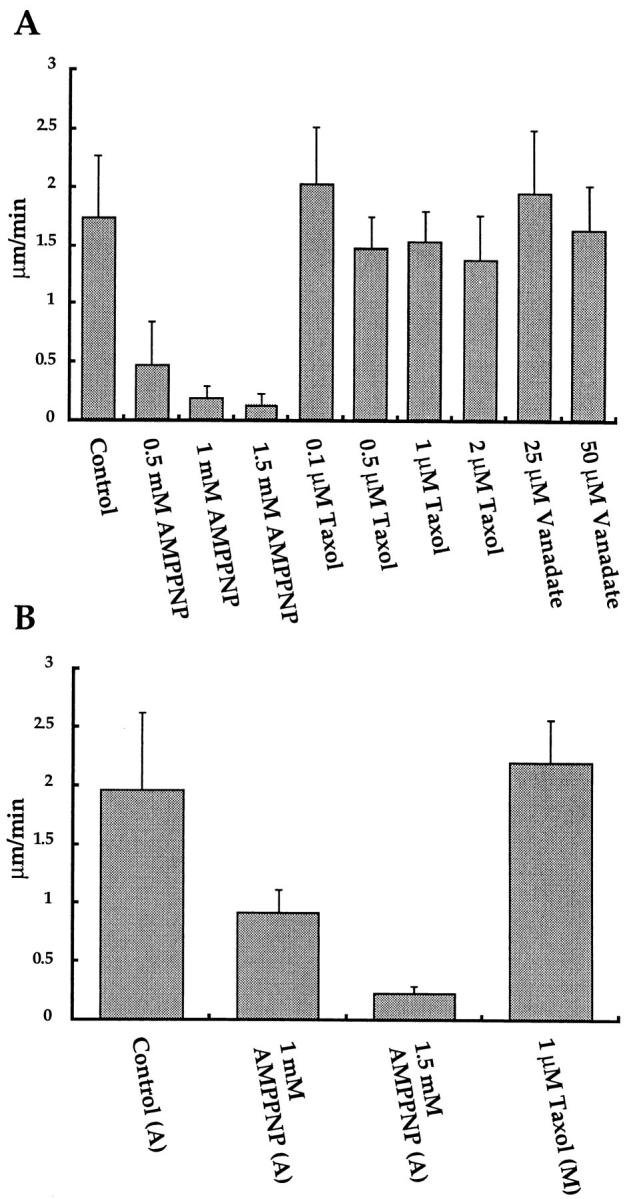
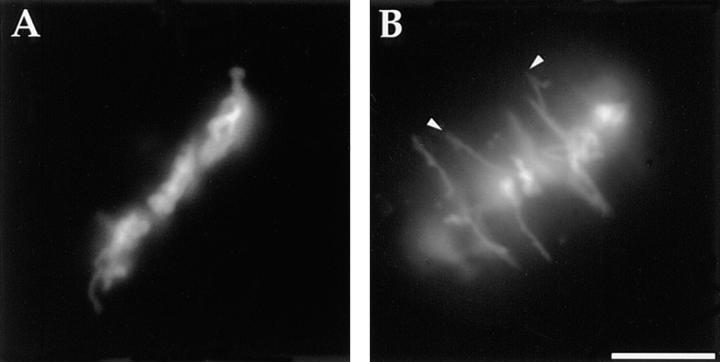
We have used local fluorescence photoactivation to mark the lattice of spindle microtubules during anaphase A in Xenopus extract spindles. We find that both poleward spindle microtubule flux and anaphase A chromosome movement occur at similar rates ( approximately 2 microm/min). This result suggests that poleward microtubule flux, coupled to microtubule depolymerization near the spindle poles, is the predominant mechanism for anaphase A in Xenopus egg extracts. In contrast, in vertebrate somatic cells a "Pacman" kinetochore mechanism, coupled to microtubule depolymerization near the kinetochore, predominates during anaphase A. Consistent with the conclusion from fluorescence photoactivation analysis, both anaphase A chromosome movement and poleward spindle microtubule flux respond similarly to pharmacological perturbations in Xenopus extracts. Furthermore, the pharmacological profile of anaphase A in Xenopus extracts differs from the previously established profile for anaphase A in vertebrate somatic cells. The difference between these profiles is consistent with poleward microtubule flux playing the predominant role in anaphase chromosome movement in Xenopus extracts, but not in vertebrate somatic cells. We discuss the possible biological implications of the existence of two distinct anaphase A mechanisms and their differential contributions to poleward chromosome movement in different cell types.
Figures
References
-
- Desai, A., and T.J. Mitchison. 1998. Preparation and characterization of caged fluorescein tubulin. Methods Enzymol. In press. - PubMed
-
- Desai, A., A.W. Murray, T.J. Mitchison, and C.E. Walczak. 1998. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. In press. - PubMed
