

A ribosomal function is necessary for efficient splicing of the T4 phage thymidylate synthase intron in vivo
- PMID: 9573049
- PMCID: PMC316773
- DOI: 10.1101/gad.12.9.1327
A ribosomal function is necessary for efficient splicing of the T4 phage thymidylate synthase intron in vivo
Abstract
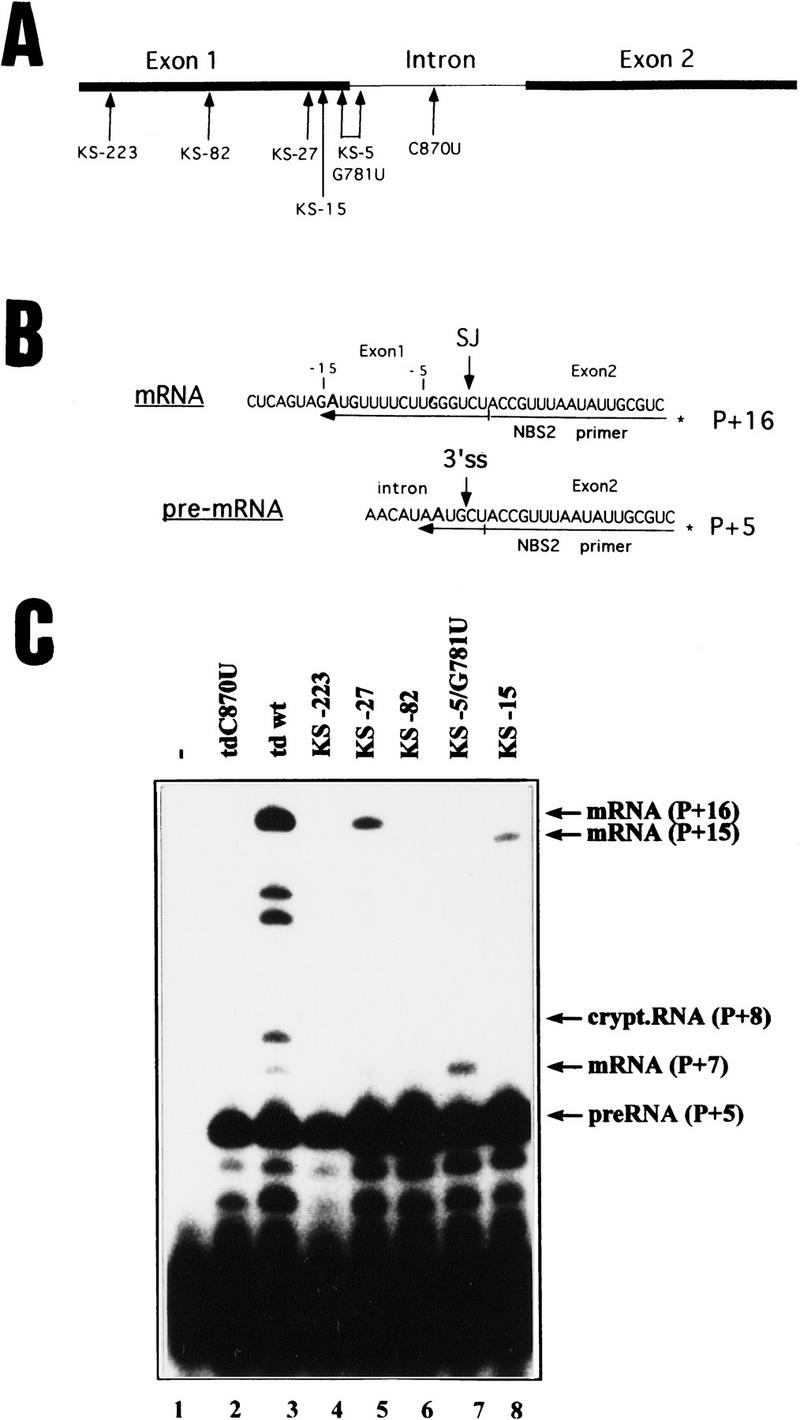
Splicing of the group I intron of the T4 thymidylate synthase (td) gene was uncoupled from translation by introducing stop codons in the upstream exon. This resulted in severe splicing deficiency in vivo. Overexpression of a UGA suppressor tRNA partially rescued splicing, suggesting that this in vitro self-splicing intron requires translation for splicing in vivo. Inhibition of translation by the antibiotics chloramphenicol and spectinomycin also resulted in splicing deficiency. Ribosomal protein S12, a protein with RNA chaperone activity, and CYT-18, a protein that stabilizes the three-dimensional structure of group I introns, efficiently rescued the stop codon mutants. We identified a region in the upstream exon that interferes with splicing. Point mutations in this region efficiently alleviate the effect of a nonsense codon. We infer from these results that the ribosome acts as an RNA chaperone to facilitate proper folding of the intron.
Figures








References
-
- Bacher G, Luetcke H, Jungnickel B, Rapoport TA, Dobberstein B. Regulation by the ribosome of the GTPase of the signal-recognition particle during protein targeting. Nature. 1996;381:248–251. - PubMed
-
- Belfort M, Moelleken A, Maley GF, Maley F. Purification and properties of T4 thymidylate synthase produced by the cloned gene in an amplification vector. J Biol Chem. 1983;258:2045–2051. - PubMed
-
- Belfort M, Chandry PS, Pedersen-Lane J. Genetic delineation of functional components of the group I intron in the phage T4 td gene. Cold Spring Harbor Symp Quant Biol. 1987;52:181–192. - PubMed
-
- Belfort M, Ehrenman K, Chandry PS. Genetic and molecular analysis of RNA splicing in Escherichia coli. Methods Enzymol. 1990;181:521–539. - PubMed
-
- Caprara MG, Lehnert V, Lambowitz AM, Westhof E. A tyrosyl-tRNA synthetase recognizes a conserved tRNA-like structural motif in the group I intron catalytic core. Cell. 1996a;87:1135–1145. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases