

Prespore-specific gene expression in Bacillus subtilis is driven by sequestration of SpoIIE phosphatase to the prespore side of the asymmetric septum
- PMID: 9573053
- PMCID: PMC316783
- DOI: 10.1101/gad.12.9.1371
Prespore-specific gene expression in Bacillus subtilis is driven by sequestration of SpoIIE phosphatase to the prespore side of the asymmetric septum
Abstract
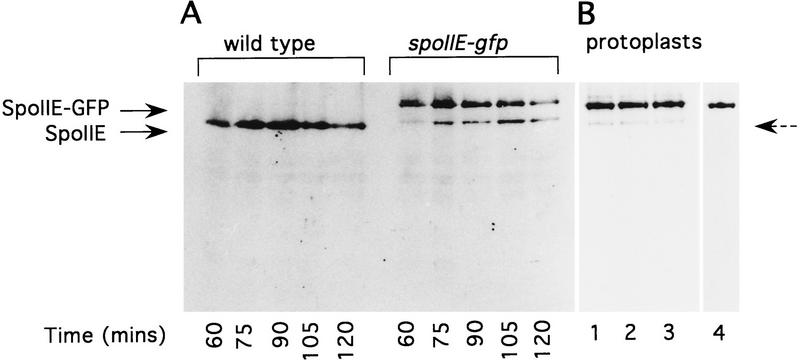
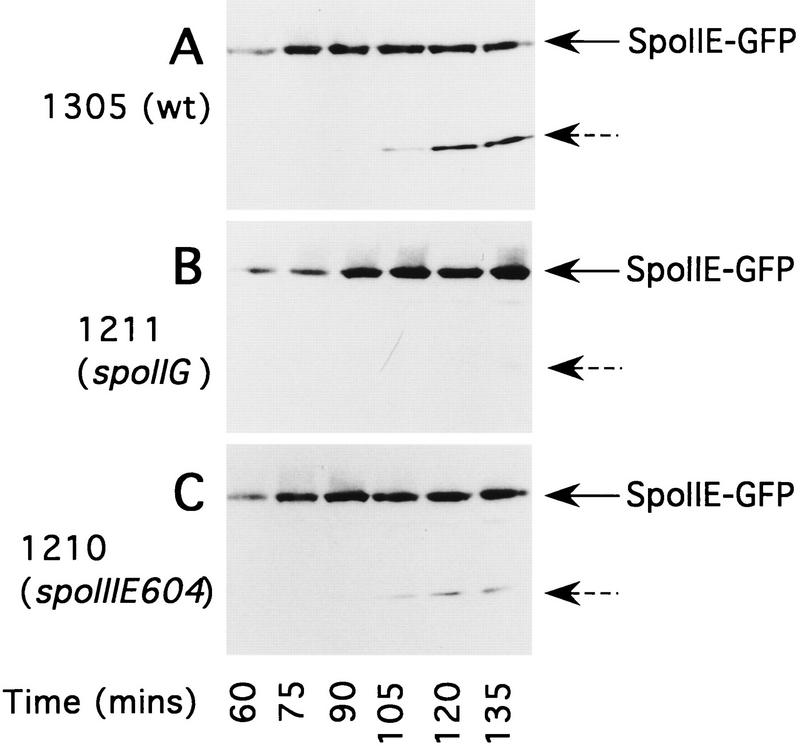
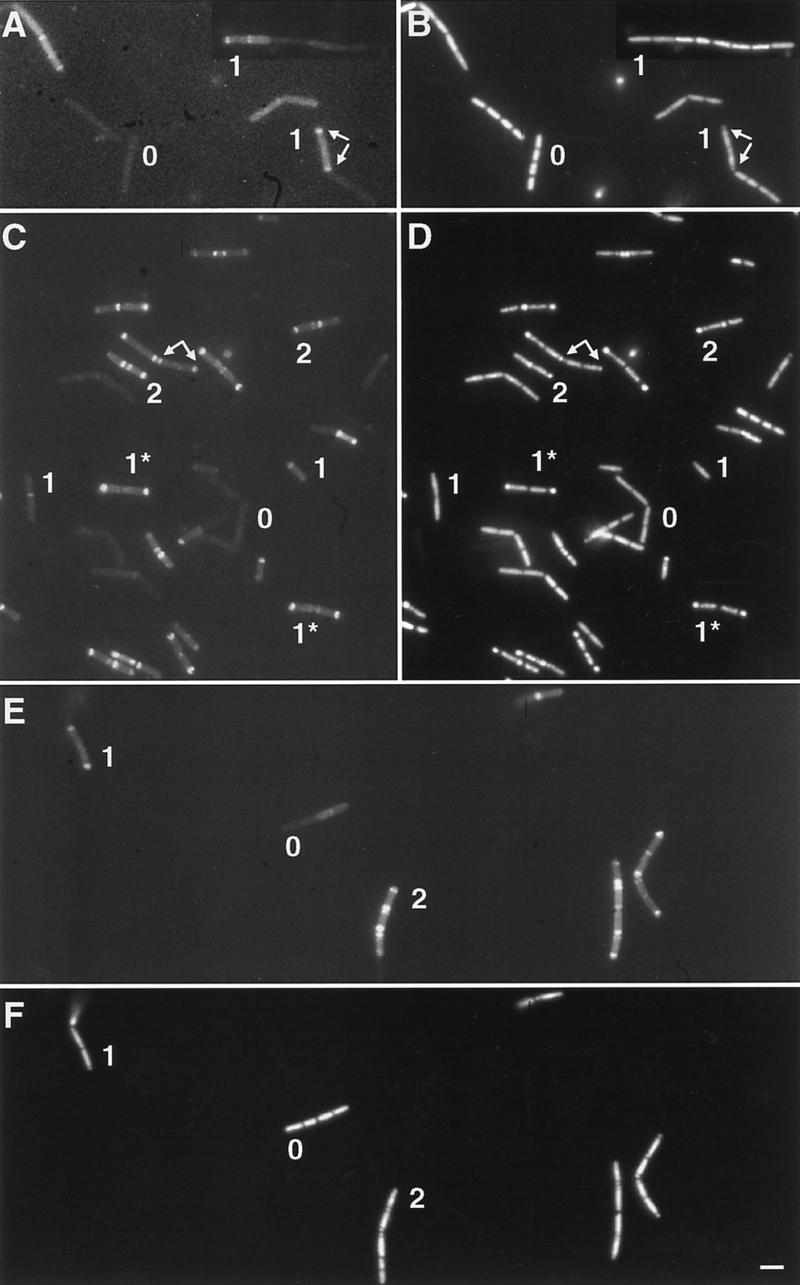
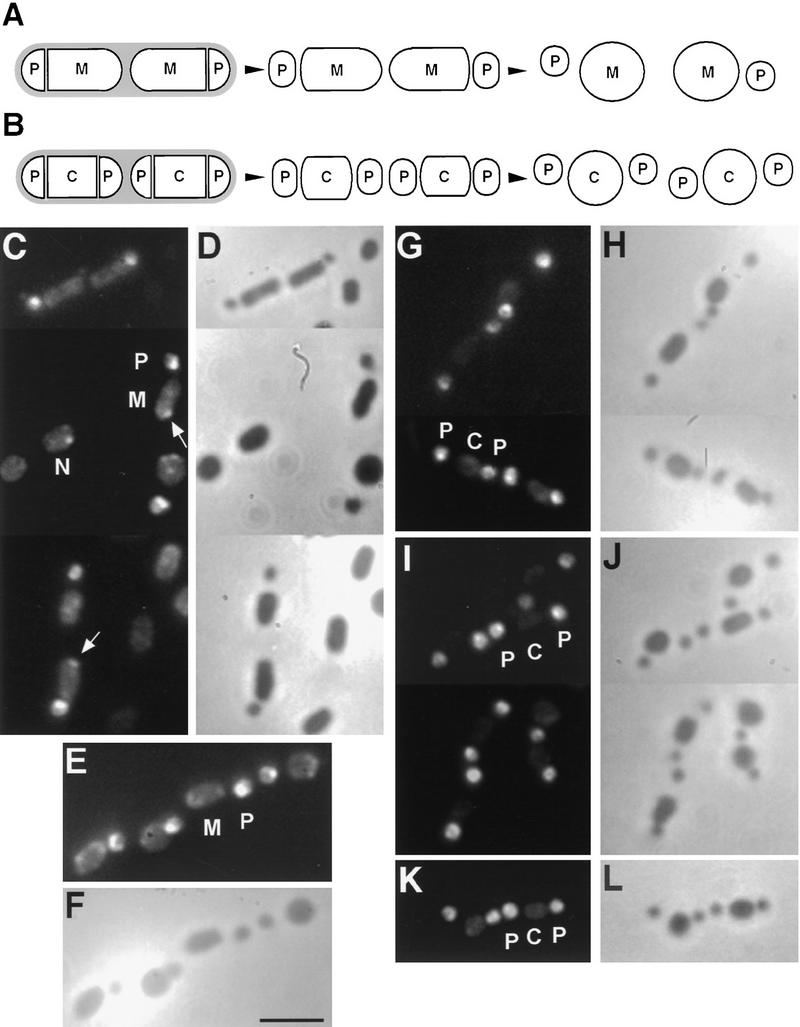
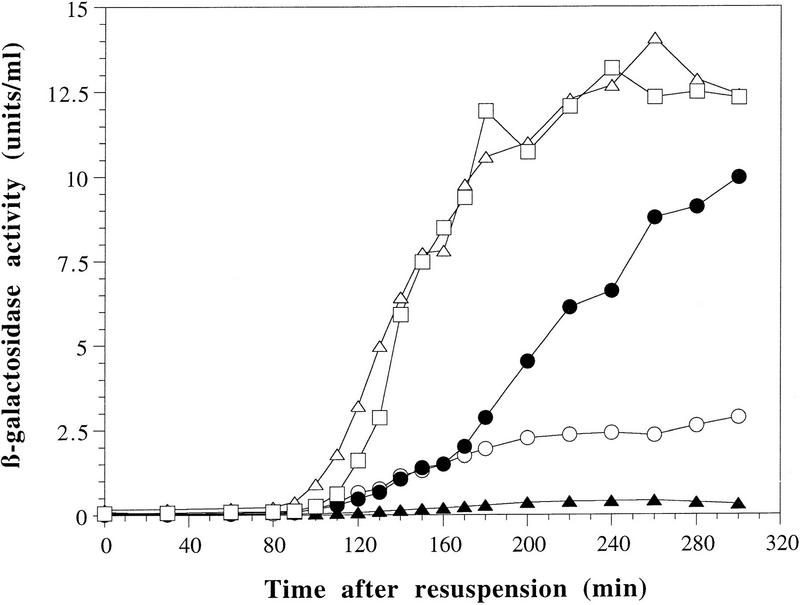
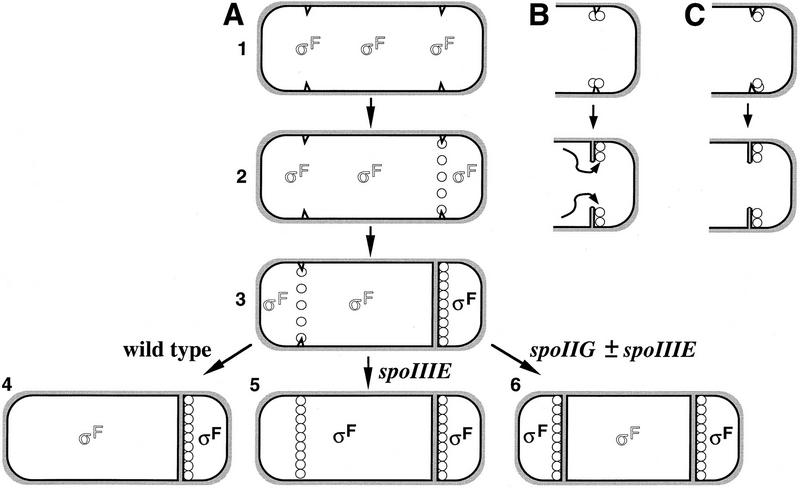
The spoIIE gene is essential for the compartment-specific activation of transcription factor sigmaF during sporulation in Bacillus subtilis. SpoIIE is a membrane protein that is targeted to the potential sites of asymmetric septation near each pole of the sporulating cell. The cytoplasmic carboxy-terminal domain of SpoIIE contains a serine phosphatase that triggers the release of sigmaF in the prespore compartment after septation. To understand how septum-located SpoIIE is activated selectively in the prespore, we examined the distribution of a SpoIIE-GFP fusion protein. We show that the polar bands of SpoIIE protein actually form sequentially and that the most prominent band develops at the pole where the prespore forms. We also show that the protein is sequestered to the prespore side of the asymmetric septum. Sequestration of SpoIIE into the prespore compartment provides a mechanism that could explain the cell specificity of sigmaF activation.
Figures
Similar articles
-
Septation, dephosphorylation, and the activation of sigmaF during sporulation in Bacillus subtilis.Genes Dev. 1999 May 1;13(9):1156-67. doi: 10.1101/gad.13.9.1156. Genes Dev. 1999. PMID: 10323866 Free PMC article.
-
Characterization of a morphological checkpoint coupling cell-specific transcription to septation in Bacillus subtilis.Mol Microbiol. 1999 Sep;33(5):1015-26. doi: 10.1046/j.1365-2958.1999.01543.x. Mol Microbiol. 1999. PMID: 10476035
-
The cell differentiation protein SpoIIE contains a regulatory site that controls its phosphatase activity in response to asymmetric septation.Mol Microbiol. 2002 Aug;45(4):1119-30. doi: 10.1046/j.1365-2958.2002.03082.x. Mol Microbiol. 2002. PMID: 12180929
-
Differential gene expression in genetically identical sister cells: the initiation of sporulation in Bacillus subtilis.Mol Microbiol. 2005 May;56(3):578-89. doi: 10.1111/j.1365-2958.2005.04594.x. Mol Microbiol. 2005. PMID: 15819616 Review.
-
Control of the cell-specificity of sigma F activity in Bacillus subtilis.Philos Trans R Soc Lond B Biol Sci. 1996 Apr 29;351(1339):537-42. doi: 10.1098/rstb.1996.0052. Philos Trans R Soc Lond B Biol Sci. 1996. PMID: 8735276 Review.
Cited by
-
Inferring Biological Mechanisms by Data-Based Mathematical Modelling: Compartment-Specific Gene Activation during Sporulation in Bacillus subtilis as a Test Case.Adv Bioinformatics. 2011;2011:124062. doi: 10.1155/2011/124062. Epub 2012 Jan 23. Adv Bioinformatics. 2011. PMID: 22312331 Free PMC article.
-
Asymmetric localization of the cell division machinery during Bacillus subtilis sporulation.Elife. 2021 May 21;10:e62204. doi: 10.7554/eLife.62204. Elife. 2021. PMID: 34018921 Free PMC article.
-
Contrasting effects of sigmaE on compartmentalization of sigmaF activity during sporulation of Bacillus subtilis.J Bacteriol. 2004 Apr;186(7):1983-90. doi: 10.1128/JB.186.7.1983-1990.2004. J Bacteriol. 2004. PMID: 15028681 Free PMC article.
-
Direct interaction between the cell division protein FtsZ and the cell differentiation protein SpoIIE.EMBO J. 2000 Apr 3;19(7):1467-75. doi: 10.1093/emboj/19.7.1467. EMBO J. 2000. PMID: 10747015 Free PMC article.
-
Mapping the bacterial cell architecture into the chromosome.Philos Trans R Soc Lond B Biol Sci. 2000 Feb 29;355(1394):179-90. doi: 10.1098/rstb.2000.0557. Philos Trans R Soc Lond B Biol Sci. 2000. PMID: 10724454 Free PMC article. Review.
References
-
- Alper S, Duncan L, Losick R. An adenosine nucleotide switch controlling the activity of a cell type-specific transcription factor in B. subtilis. Cell. 1994;77:195–205. - PubMed
-
- Arigoni F, Pogliano K, Webb CD, Stragier P, Losick R. Localization of protein implicated in establishment of cell type to sites of asymmetric division. Science. 1995;270:637–640. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources