

Ligand induction of retinoic acid receptors alters an acute infection by murine cytomegalovirus
- PMID: 9573222
- PMCID: PMC109973
- DOI: 10.1128/JVI.72.6.4589-4600.1998
Ligand induction of retinoic acid receptors alters an acute infection by murine cytomegalovirus
Abstract
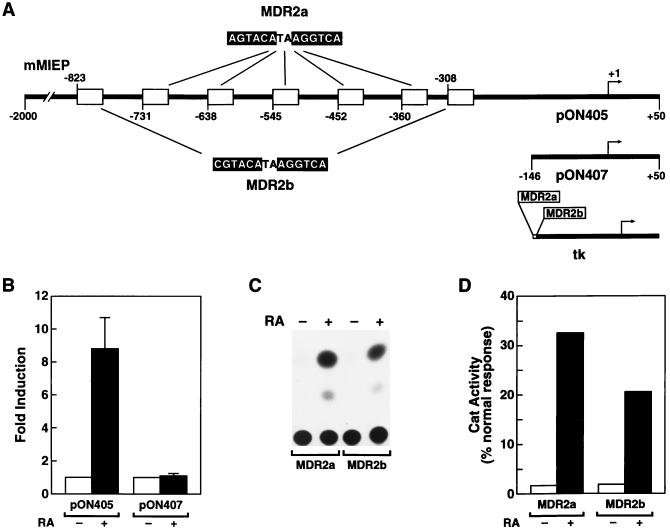
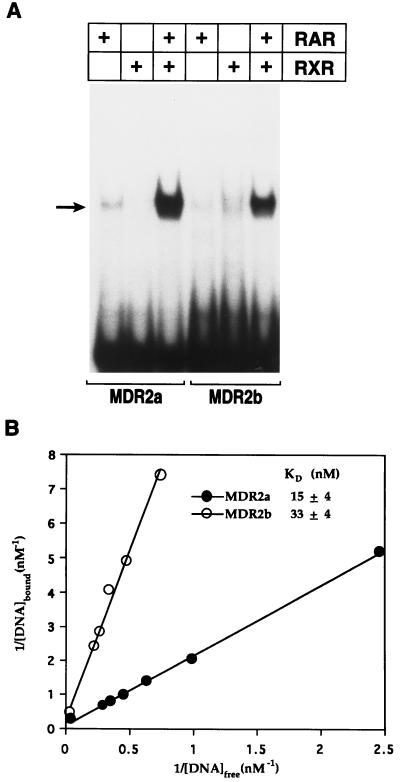
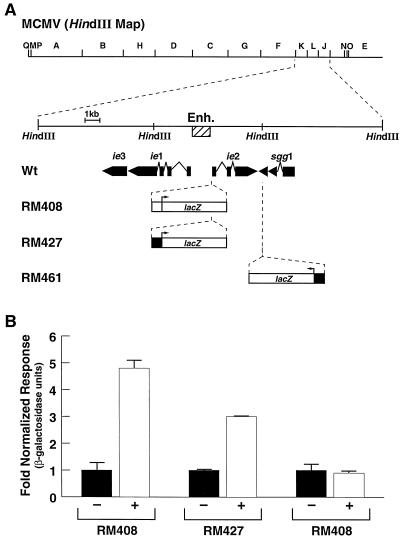
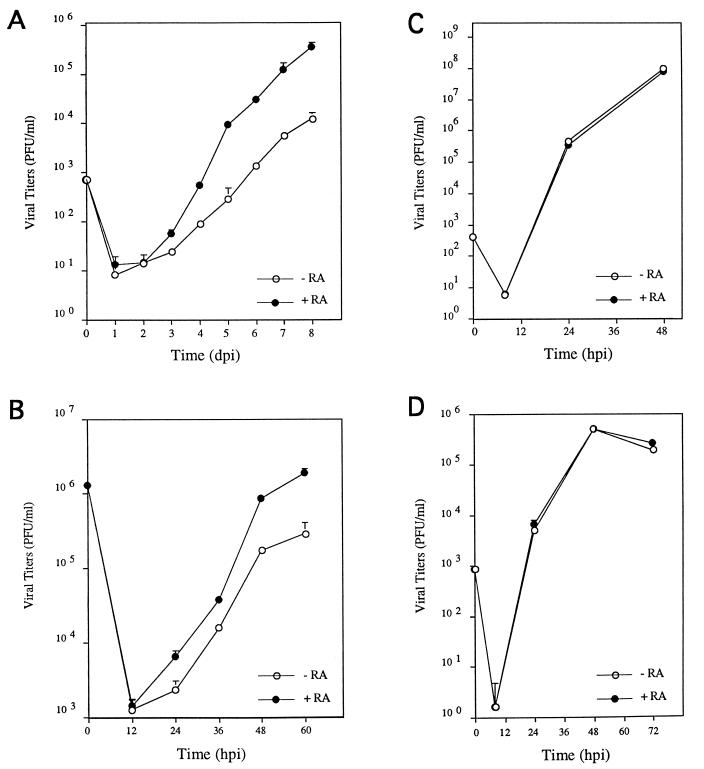
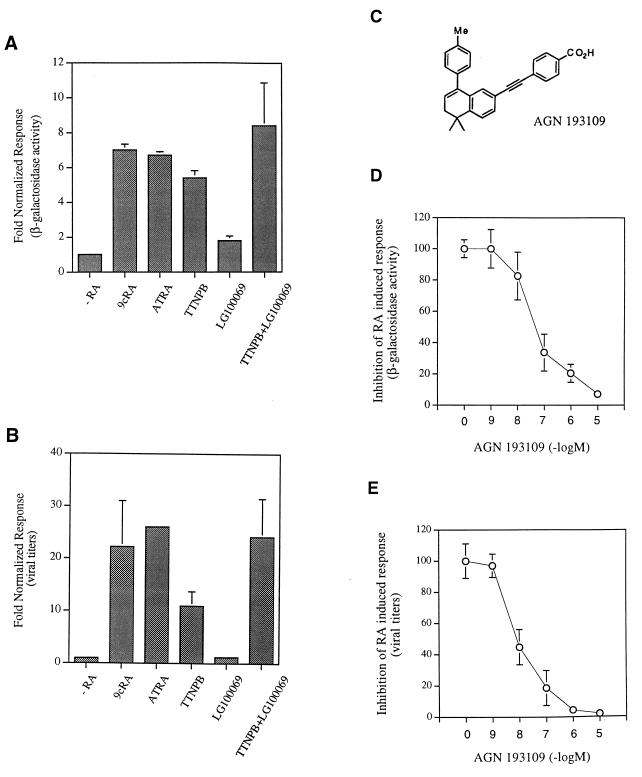
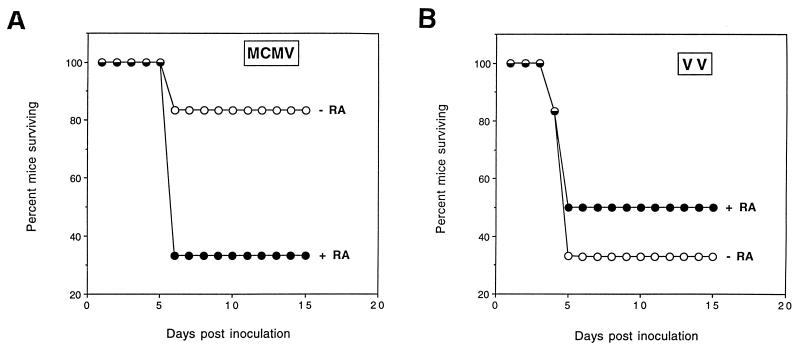
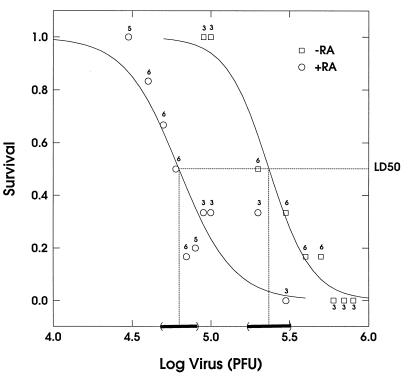
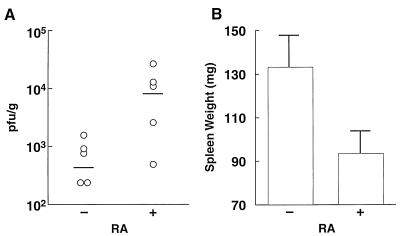
Here we report that administration of retinoids can alter the outcome of an acute murine cytomegalovirus (MCMV) infection. We show that a crucial viral control element, the major immediate-early enhancer, can be activated by retinoic acid (RA) via multiple RA-responsive elements (DR2) that bind retinoid X receptor-retinoic acid receptor (RAR) heterodimers with apparent dissociation constants ranging from 15 to 33 nM. Viral growth is dramatically increased upon RA treatment of infected tissue culture cells. Using synthetic retinoid receptor-specific agonists and antagonists, we provide evidence that RAR activation in cells is required for mediating the response of MCMV to RA. Oral administration of RA to infected immunocompetent mice selectively exacerbates an infection by MCMV, while cotreatment with an RAR antagonist protects against the adverse effects of RA on MCMV infection. In conclusion, these chemical genetic experiments provide evidence that an RAR-mediated pathway can modulate in vitro and in vivo infections by MCMV.
Figures
References
-
- Achkar C C, Bentel J M, Boylan J F, Scher H I, Gudas L J, Miller W H J. Differences in the pharmacokinetic properties of orally administered all-trans-retinoic acid and 9-cis-retinoic acid in the plasma of nude mice. Drug Metab Dispos. 1994;22:451–458. - PubMed
-
- Agarwal C, Chandraratna R A S, Johnson A T, Rorke E A, Eckert R L. AGN 193109 is a highly effective antagonist of retinoid action in human ectocervical epithelial cells. J Biol Chem. 1996;271:12209–12212. - PubMed
-
- Angulo A, Ghazal P. Regulation of human cytomegalovirus by retinoic acid. Scand J Infect Dis. 1995;99:113–115. - PubMed
-
- Angulo, A., and P. Ghazal. Unpublished data.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
