

Rep*: a viral element that can partially replace the origin of plasmid DNA synthesis of Epstein-Barr virus
- PMID: 9573229
- PMCID: PMC109986
- DOI: 10.1128/JVI.72.6.4657-4666.1998
Rep*: a viral element that can partially replace the origin of plasmid DNA synthesis of Epstein-Barr virus
Abstract
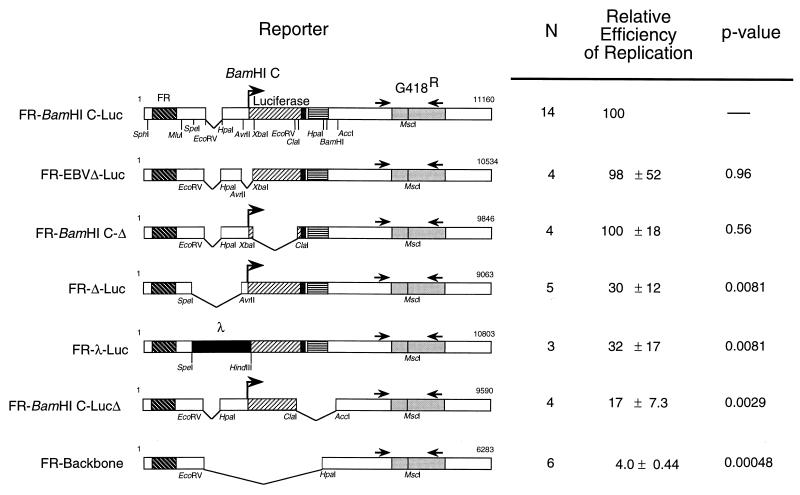
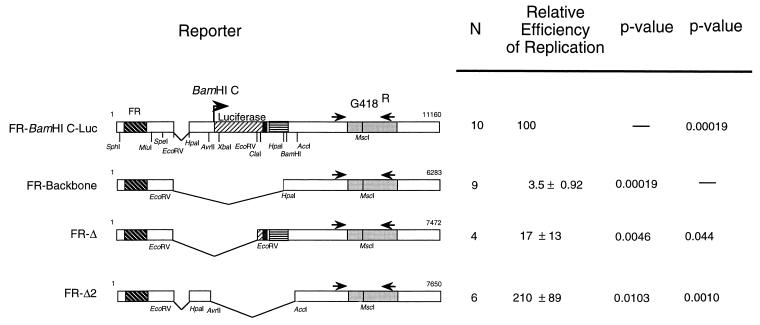
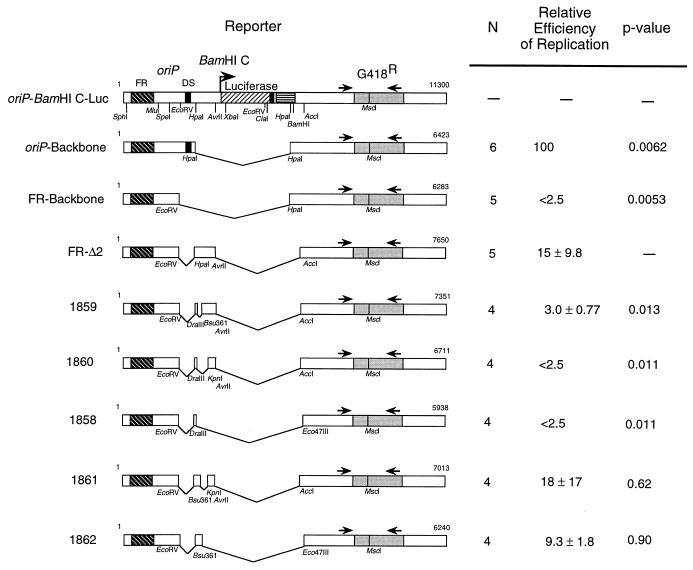
Replication of the Epstein-Barr viral (EBV) genome occurs once per cell cycle during latent infection. Similarly, plasmids containing EBV's plasmid origin of replication, oriP, are replicated once per cell cycle. Replication from oriP requires EBV nuclear antigen 1 (EBNA-1) in trans; however, its contributions to this replication are unknown. oriP contains 24 EBNA-1 binding sites; 20 are located within the family of repeats, and 4 are found within the dyad symmetry element. The site of initiation of DNA replication within oriP is at or near the dyad symmetry element. We have identified a plasmid that contains the family of repeats but lacks the dyad symmetry element whose replication can be detected for a limited number of cell cycles. The detection of short-term replication of this plasmid requires EBNA-1 and can be inhibited by a dominant-negative inhibitor of EBNA-1. We have identified two regions within this plasmid which can independently contribute to this replication in the absence of the dyad symmetry element of oriP. One region contains native EBV sequences within the BamHI C fragment of the B95-8 genome of EBV; the other contains sequences within the simian virus 40 genome. We have mapped the region contributing to replication within the EBV sequences to a 298-bp fragment, Rep*. Plasmids which contain three copies of Rep* plus the family of repeats support replication more efficiently than those with one copy, consistent with a stochastic model for the initiation of DNA synthesis. Plasmids with three copies of Rep* also support long-term replication in the presence of EBNA-1. These observations together indicate that the latent origin of replication of EBV is more complex than formerly appreciated; it is a multicomponent origin of which the dyad symmetry element is one efficient component. The experimental approach described here could be used to identify eukaryotic sequences which mediate DNA synthesis, albeit inefficiently.
Figures

Similar articles
-
Plasmid maintenance of derivatives of oriP of Epstein-Barr virus.J Virol. 1995 Feb;69(2):1280-3. doi: 10.1128/JVI.69.2.1280-1283.1995. J Virol. 1995. PMID: 7815506 Free PMC article.
-
The Epstein-Barr virus origin of plasmid replication, oriP, contains both the initiation and termination sites of DNA replication.Cell. 1989 Aug 11;58(3):527-35. doi: 10.1016/0092-8674(89)90433-9. Cell. 1989. PMID: 2547525
-
The plasmid replicon of EBV consists of multiple cis-acting elements that facilitate DNA synthesis by the cell and a viral maintenance element.EMBO J. 1998 Nov 2;17(21):6394-403. doi: 10.1093/emboj/17.21.6394. EMBO J. 1998. PMID: 9799247 Free PMC article.
-
Replication licensing of the EBV oriP minichromosome.Curr Top Microbiol Immunol. 2001;258:13-33. doi: 10.1007/978-3-642-56515-1_2. Curr Top Microbiol Immunol. 2001. PMID: 11443858 Review.
-
Latent and lytic Epstein-Barr virus replication strategies.Rev Med Virol. 2005 Jan-Feb;15(1):3-15. doi: 10.1002/rmv.441. Rev Med Virol. 2005. PMID: 15386591 Review.
Cited by
-
Replication of Epstein-Barr viral DNA.Cold Spring Harb Perspect Biol. 2013 Jan 1;5(1):a013029. doi: 10.1101/cshperspect.a013029. Cold Spring Harb Perspect Biol. 2013. PMID: 23284049 Free PMC article. Review.
-
The coupling of synthesis and partitioning of EBV's plasmid replicon is revealed in live cells.EMBO J. 2007 Oct 3;26(19):4252-62. doi: 10.1038/sj.emboj.7601853. Epub 2007 Sep 13. EMBO J. 2007. PMID: 17853891 Free PMC article.
-
Establishment of an oriP replicon is dependent upon an infrequent, epigenetic event.Mol Cell Biol. 2001 Jul;21(13):4149-61. doi: 10.1128/MCB.21.13.4149-4161.2001. Mol Cell Biol. 2001. PMID: 11390644 Free PMC article.
-
The plasmid replicon of Epstein-Barr virus: mechanistic insights into efficient, licensed, extrachromosomal replication in human cells.Plasmid. 2007 Jul;58(1):1-12. doi: 10.1016/j.plasmid.2007.01.003. Epub 2007 Mar 9. Plasmid. 2007. PMID: 17350094 Free PMC article. Review.
-
The latent origin of replication of Epstein-Barr virus directs viral genomes to active regions of the nucleus.J Virol. 2010 Mar;84(5):2533-46. doi: 10.1128/JVI.01909-09. Epub 2009 Dec 23. J Virol. 2010. PMID: 20032186 Free PMC article.
References
-
- Baer R, Bankier A T, Biggin M D, Deininger P L, Farrell P J, Gibson T J, Hatfull G, Hudson G S, Satchwell S C, Séguin C, Tuffnell P S, Barrell B G. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature. 1984;310:207–211. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
