

Vpr stimulates viral expression and induces cell killing in human immunodeficiency virus type 1-infected dividing Jurkat T cells
- PMID: 9573232
- PMCID: PMC109992
- DOI: 10.1128/JVI.72.6.4686-4693.1998
Vpr stimulates viral expression and induces cell killing in human immunodeficiency virus type 1-infected dividing Jurkat T cells
Abstract
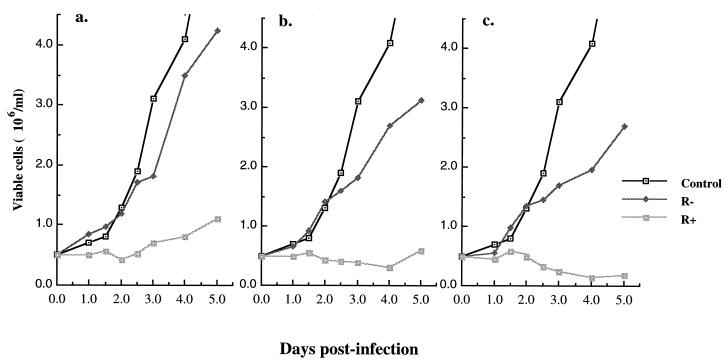
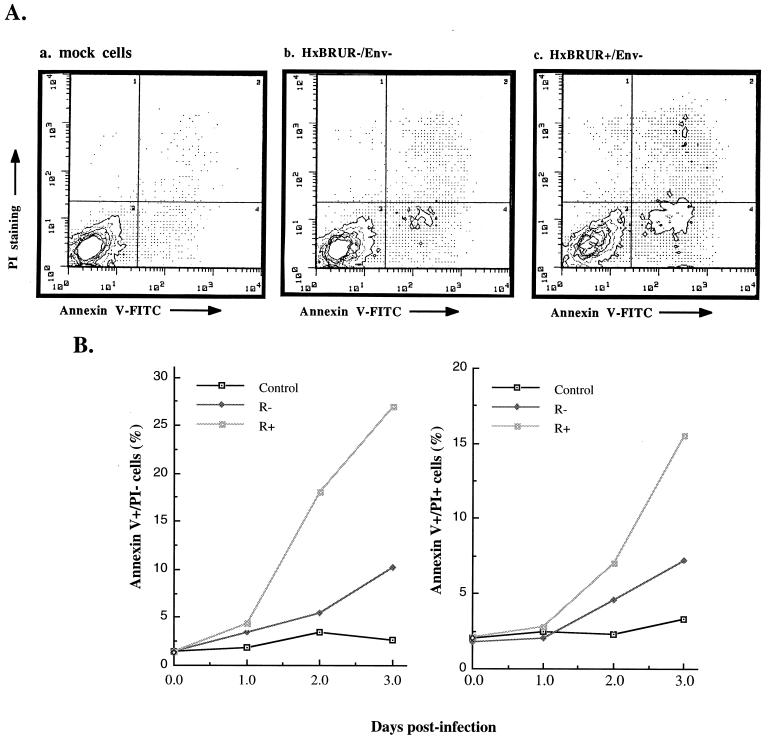
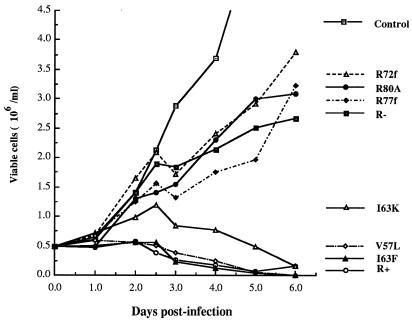
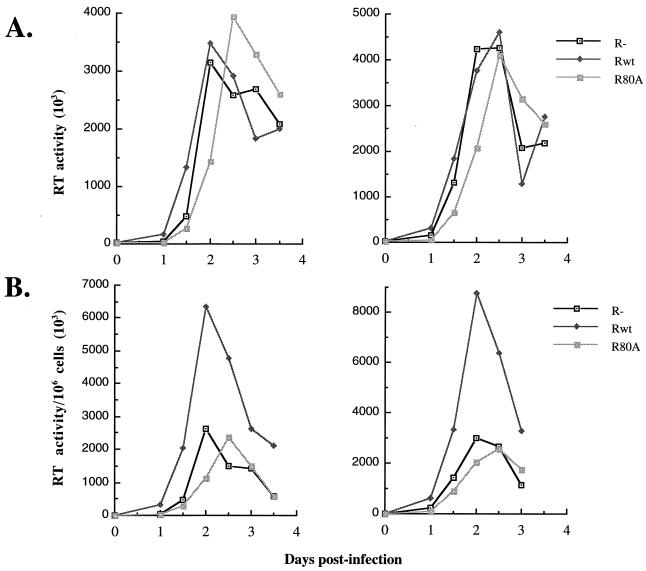
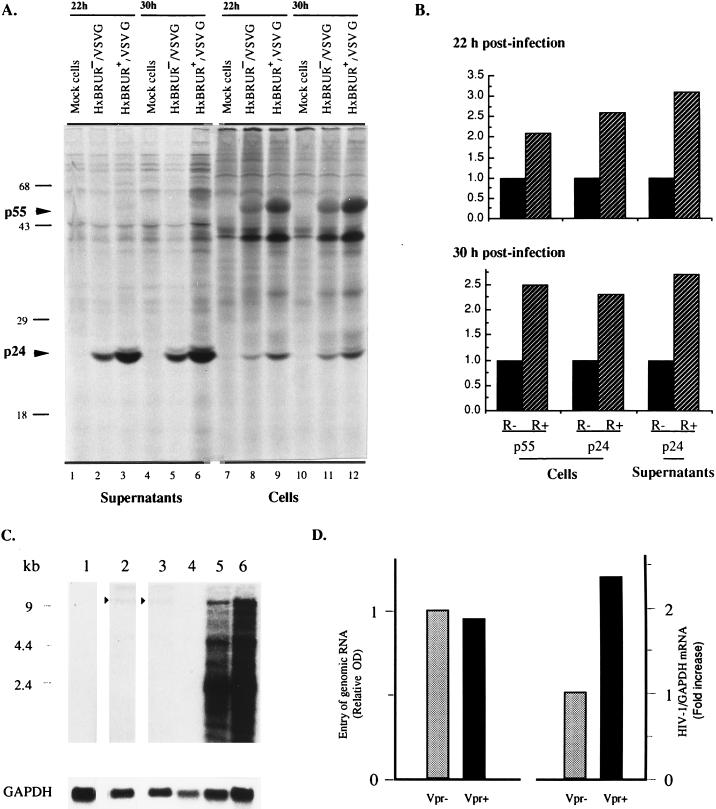
In this study we investigated the effects of Vpr during human immunodeficiency virus (HIV) infection of proliferating Jurkat T cells by using a vesicular stomatitis virus envelope G glycoprotein pseudotyped HIV superinfection system. We observe that the expression of Vpr results in a severe reduction in the life span of HIV type 1 (HIV-1)-infected dividing T cells in culture. In agreement with a recent report (S. A. Stewart, B. Poon, J. B. M. Jowett, and I. S. Chen, J. Virol. 71:5579-5592, 1997), we show that events characteristic of apoptotic cell death are involved in the Vpr-mediated cytopathic effects. Our results also show that infection with viruses expressing the wild-type vpr gene results in an increase in viral gene expression and production. Interestingly, the effects of Vpr on cell viability and on viral gene expression both correlate with the ability of the protein to induce a cell cycle arrest in the G2/M phase. Mutagenesis analyses show that the C terminus of Vpr is essential for these biological activities. Although the role of Vpr is currently associated with the infection of nondividing cells, our results suggest that Vpr can also directly increase viral replication in vivo in infected dividing T cells. Furthermore, these in vitro observations suggest that Vpr-mediated cytotoxic effects could contribute to the CD4+ depletion associated with AIDS progression.
Figures
References
-
- Agostini I, Navarro J M, Rey F, Bouhamdan M, Spire B, Vigne R, Sire J. The human immunodeficiency virus type 1 Vpr transactivator: cooperation with promoter-bound activator domains and binding to TFIIB. J Mol Biol. 1996;261:599–606. - PubMed
-
- Ayyavoo V, Mahboubi A, Mahalingam S, Ramalingam R, Kudchodkar S, Williams W V, Green D R, Weiner D B. HIV-1 Vpr suppresses immune activation and apoptosis through regulation of nuclear factor κB. Nat Med. 1997;3:1117–1123. - PubMed
-
- Balliet J W, Kolson D L, Eiger G, Kim F M, McGann K A, Srinivasan A, Collman R. Distinct effects in primary macrophages and lymphocytes of the human immunodeficiency virus type 1 accessory genes vpr, vpu, and nef: mutational analysis of a primary HIV-1 isolate. Virology. 1994;200:623–631. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
