

Long-term evolution of the hypervariable region of hepatitis C virus in a common-source-infected cohort
- PMID: 9573256
- PMCID: PMC110045
- DOI: 10.1128/JVI.72.6.4893-4905.1998
Long-term evolution of the hypervariable region of hepatitis C virus in a common-source-infected cohort
Abstract
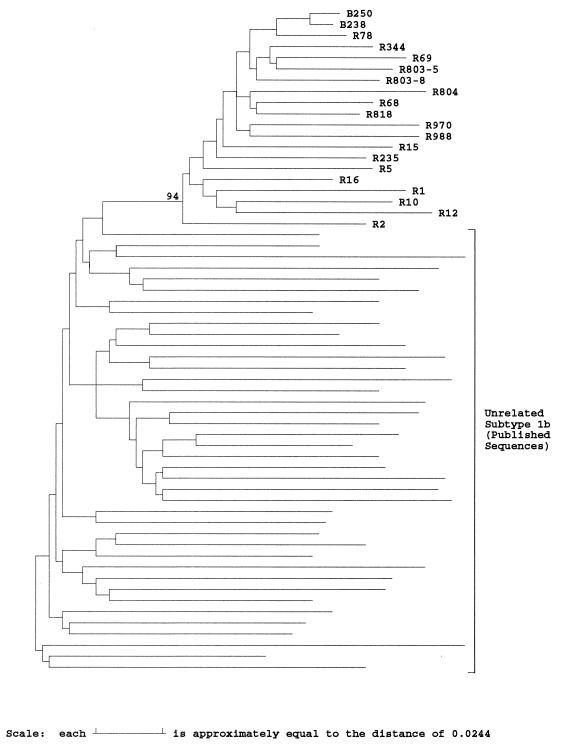
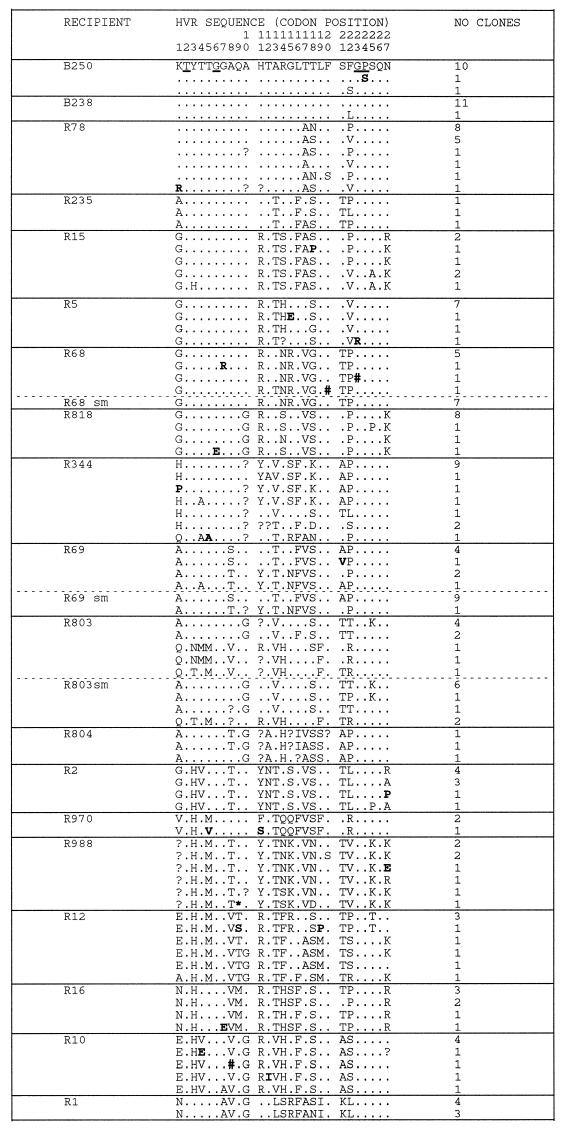
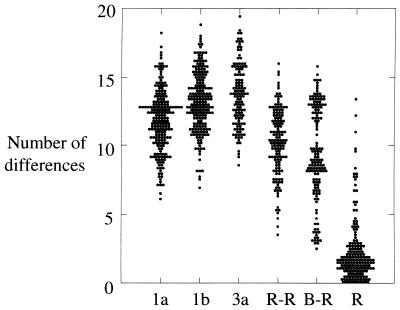
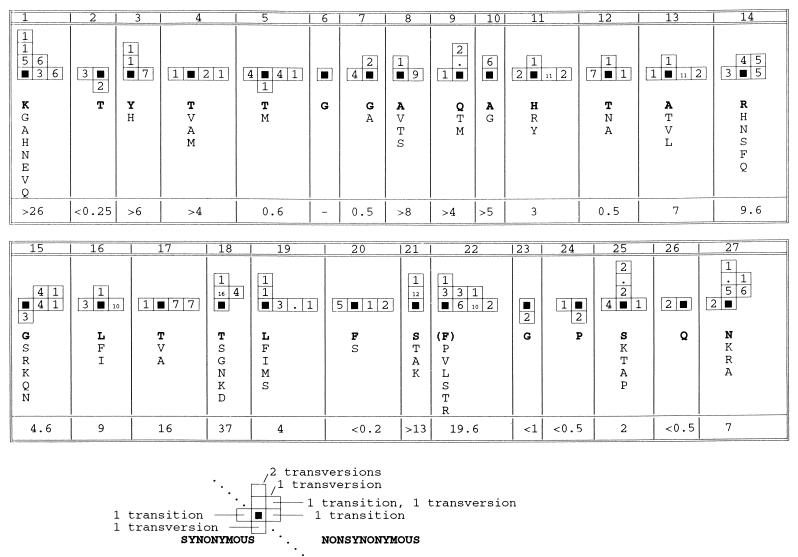
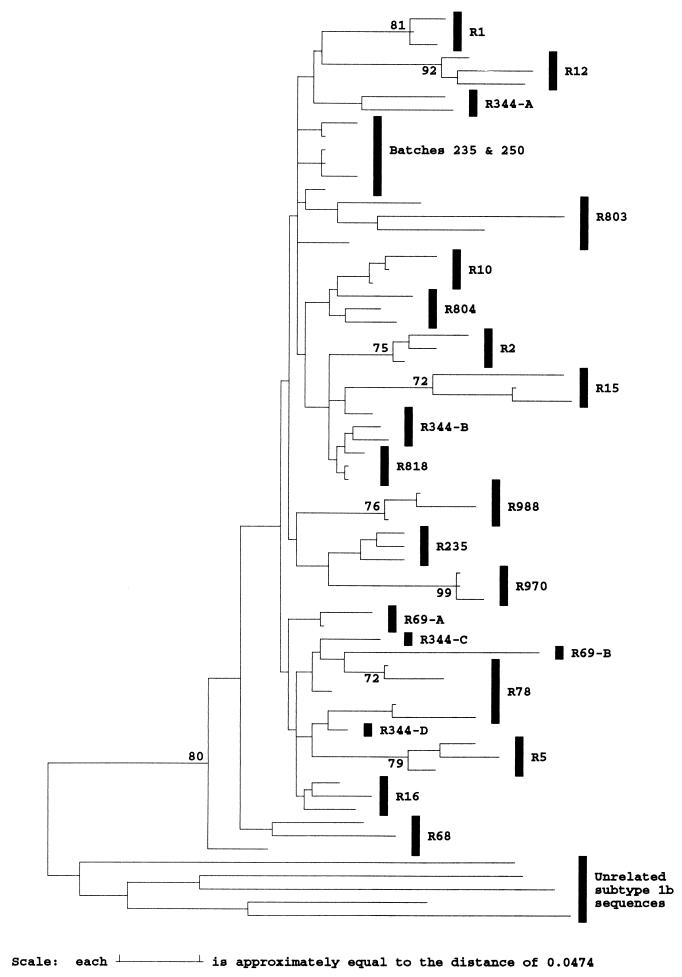
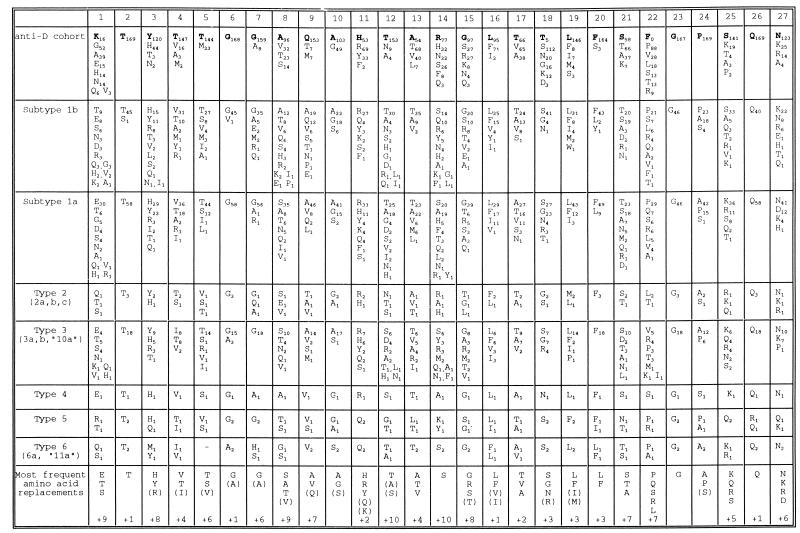
The long-term evolution of the hepatitis C virus hypervariable region (HVR) and flanking regions of the E1 and E2 envelope proteins have been studied in a cohort of women infected from a common source of anti-D immunoglobulin. Whereas virus sequences in the infectious source were relatively homogeneous, distinct HVR variants were observed in each anti-D recipient, indicating that this region can evolve in multiple directions from the same point. Where HVR variants with dissimilar sequences were present in a single individual, the frequency of synonymous substitution in the flanking regions suggested that the lineages diverged more than a decade previously. Even where a single major HVR variant was present in an infected individual, this lineage was usually several years old. Multiple lineages can therefore coexist during long periods of chronic infection without replacement. The characteristics of amino acid substitution in the HVR were not consistent with the random accumulation of mutations and imply that amino acid replacement in the HVR was strongly constrained. Another variable region of E2 centered on codon 60 shows similar constraints, while HVR2 was relatively unconstrained. Several of these features are difficult to explain if a neutralizing immune response against the HVR is the only selective force operating on E2. The impact of PCR artifacts such as nucleotide misincorporation and the shuffling of dissimilar templates is discussed.
Figures
References
-
- Allander T, Beyene A, Jacobson S H, Grillner L, Persson M A A. Patients infected with the same hepatitis C virus strain display different kinetics of the isolate-specific antibody response. J Infect Dis. 1997;175:26–31. - PubMed
-
- Driesel G, Wirth D, Stark K, Baumgarten R, Sucker U, Schreier E. Hepatitis C virus (HCV) genotype distribution in German isolates: studies on the sequence variability in the e2 and NS5 region. Arch Virol. 1994;139:379–388. - PubMed
-
- Enomoto N, Sakuma I, Asahina Y, Kurosaki M, Murakami T, Yamamoto C, Izumi N, Marumo F, Sato C. Comparison of full-length sequences of interferon-sensitive and resistant hepatitis C virus 1b—sensitivity to interferon is conferred by amino acid substitutions in the NS5a region. J Clin Invest. 1995;96:224–230. - PMC - PubMed
-
- Esteban J I, Gomez J, Martell M, Cabot B, Quer J, Camps J, Gonzalez A, Otero T, Moya A, Esteban R, Guardia J. Transmission of hepatitis C virus by a cardiac surgeon. N Engl J Med. 1996;334:555–560. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
