

Human cytomegalovirus oriLyt sequence requirements
- PMID: 9573268
- PMCID: PMC110061
- DOI: 10.1128/JVI.72.6.4989-4996.1998
Human cytomegalovirus oriLyt sequence requirements
Abstract
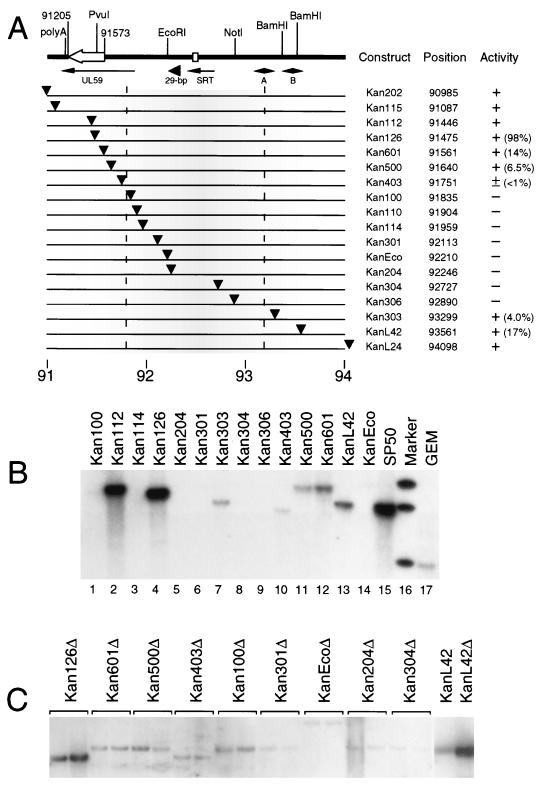
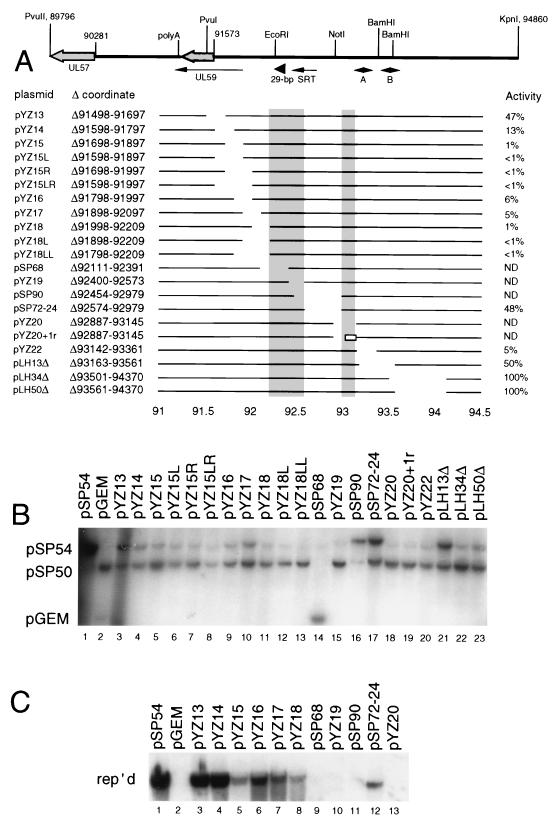
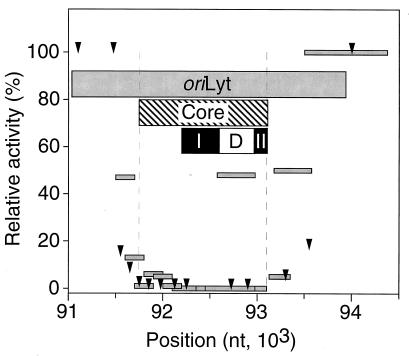
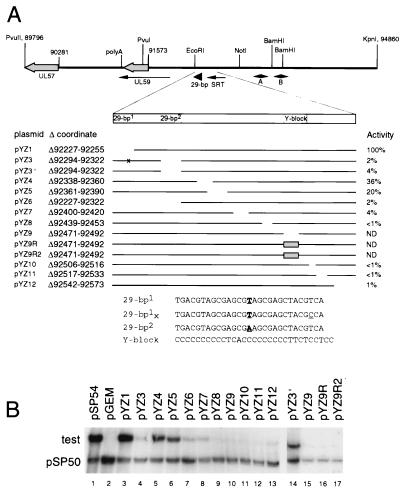
The mechanisms of action and regulation of the human cytomegalovirus (HCMV) lytic-phase DNA replicator, oriLyt, which spans more than 2 kbp in a structurally complex region near the middle of the unique long region (UL), are not understood. Because oriLyt is thought to be essential for promoting initiation of lytic DNA synthesis and may participate in regulating the switch between lytic and latent phases, we undertook a mutational study to better define its sequence requirements. Kanr gene cassette insertions located an oriLyt core region between nucleotides (nt) 91751 and 93299 that is necessary but not sufficient for replicator activity in transient assays. In contrast, insertions into auxiliary regions flanking either side of this core-also required for significant replicator activity-had little effect. To search for essential components within the core region, we made a series of overlapping, roughly 200-bp deletions, and qualitatively and quantitatively assessed the abilities of the resulting constructs to mediate replication. All but one of these deletions produced a significant (i.e., greater than twofold) loss of activity, arguing that sequences across this entire region contribute to replicator function. However, two particularly critical segments separated by a dispensable region, here called essential regions I and II, were identified. Within essential region I, which overlaps the previously identified early transcript SRT, two adjacent but nonoverlapping, roughly 200-bp deletions abolished detectable replication. No single element or motif from the left half of essential region I was found to be essential. Thus, essential region I probably promotes replication through the cooperation of multiple elements. However, four small deletions in the right half of essential region I, which included or lay adjacent to the conserved 31-nt oligopyrimidine tract (referred to as the Y block), abolished or virtually abolished oriLyt activity. Together, these results identify candidate oriLyt sequences within which molecular interactions essential for initiation of oriLyt-mediated DNA synthesis are likely to occur.
Figures
Similar articles
-
Analysis of human cytomegalovirus oriLyt sequence requirements in the context of the viral genome.J Virol. 2005 Mar;79(6):3615-26. doi: 10.1128/JVI.79.6.3615-3626.2005. J Virol. 2005. PMID: 15731256 Free PMC article.
-
The variable 3' ends of a human cytomegalovirus oriLyt transcript (SRT) overlap an essential, conserved replicator element.J Virol. 1996 Aug;70(8):5272-81. doi: 10.1128/JVI.70.8.5272-5281.1996. J Virol. 1996. PMID: 8764037 Free PMC article.
-
Evidence that the UL84 gene product of human cytomegalovirus is essential for promoting oriLyt-dependent DNA replication and formation of replication compartments in cotransfection assays.J Virol. 1996 Nov;70(11):7398-413. doi: 10.1128/JVI.70.11.7398-7413.1996. J Virol. 1996. PMID: 8892858 Free PMC article.
-
Nuts and bolts of human cytomegalovirus lytic DNA replication.Curr Top Microbiol Immunol. 2008;325:153-66. doi: 10.1007/978-3-540-77349-8_9. Curr Top Microbiol Immunol. 2008. PMID: 18637505 Review.
-
[The mechanisms of human cytomegalovirus DNA replication].Nihon Rinsho. 1998 Jan;56(1):36-43. Nihon Rinsho. 1998. PMID: 9465662 Review. Japanese.
Cited by
-
Analysis of human cytomegalovirus oriLyt sequence requirements in the context of the viral genome.J Virol. 2005 Mar;79(6):3615-26. doi: 10.1128/JVI.79.6.3615-3626.2005. J Virol. 2005. PMID: 15731256 Free PMC article.
-
Complete sequence and genomic analysis of rhesus cytomegalovirus.J Virol. 2003 Jun;77(12):6620-36. doi: 10.1128/jvi.77.12.6620-6636.2003. J Virol. 2003. PMID: 12767982 Free PMC article.
-
Initiation of lytic DNA replication in Epstein-Barr virus: search for a common family mechanism.Future Virol. 2010 Jan;5(1):65-83. doi: 10.2217/fvl.09.69. Future Virol. 2010. PMID: 22468146 Free PMC article.
-
Nonrandom clusters of palindromes in herpesvirus genomes.J Comput Biol. 2005 Apr;12(3):331-54. doi: 10.1089/cmb.2005.12.331. J Comput Biol. 2005. PMID: 15857246 Free PMC article. Review.
-
pUL34 binding near the human cytomegalovirus origin of lytic replication enhances DNA replication and viral growth.Virology. 2018 May;518:414-422. doi: 10.1016/j.virol.2018.03.017. Epub 2018 Apr 5. Virology. 2018. PMID: 29626748 Free PMC article.
References
-
- Alford C A, Britt W J. Cytomegalovirus. In: Roizman B, Whitley R I, Lopez C, editors. The human herpesviruses. 1st ed. New York, NY: Raven Press; 1993. pp. 227–255.
-
- Anders, D. G., M. B. Coy, and Y. Zhu. Unpublished results.
-
- Anders D G, McCue L A. The human cytomegalovirus genes and proteins required for DNA synthesis. Intervirology. 1996;39:378–388. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
