

From estrogen to androgen receptor: a new pathway for sex hormones in prostate
- PMID: 9576916
- PMCID: PMC20411
- DOI: 10.1073/pnas.95.10.5527
From estrogen to androgen receptor: a new pathway for sex hormones in prostate
Abstract
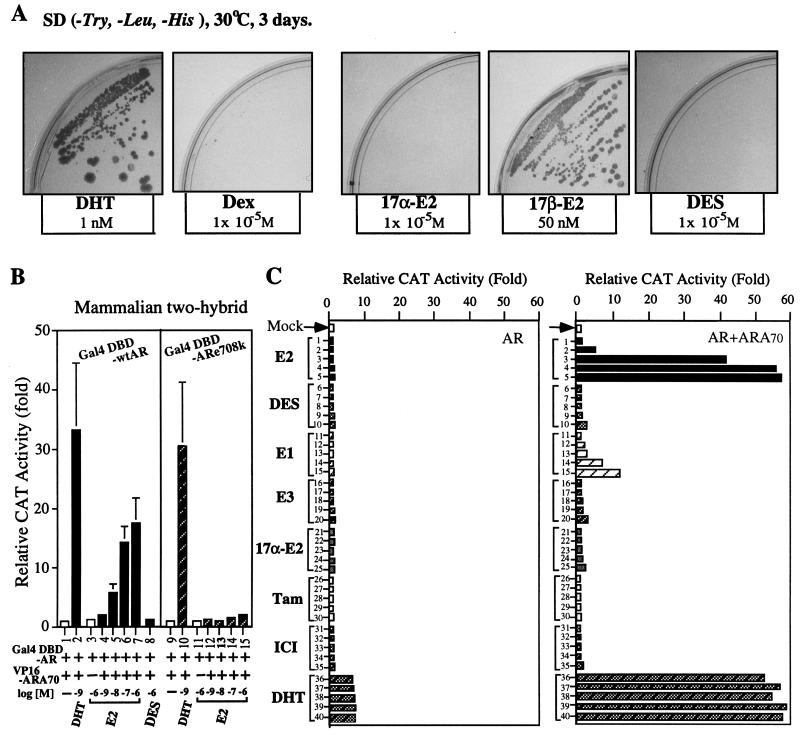
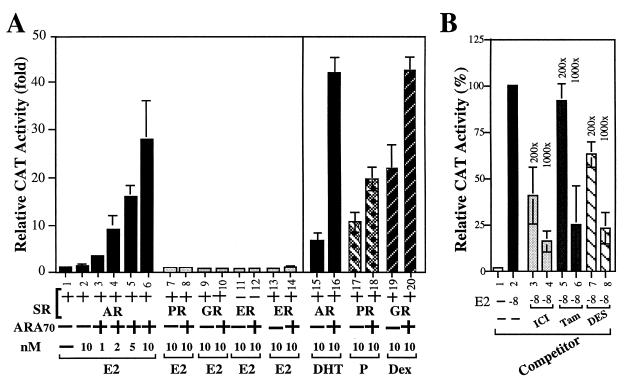
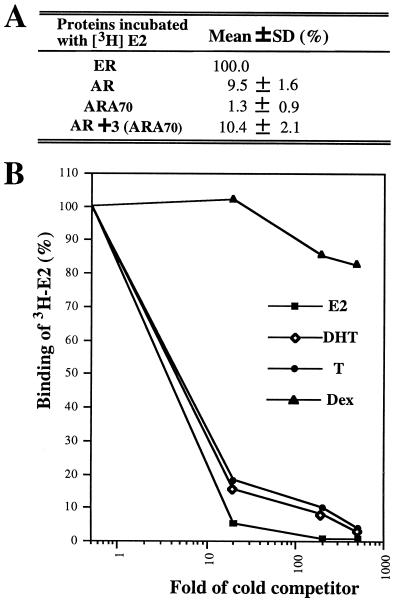
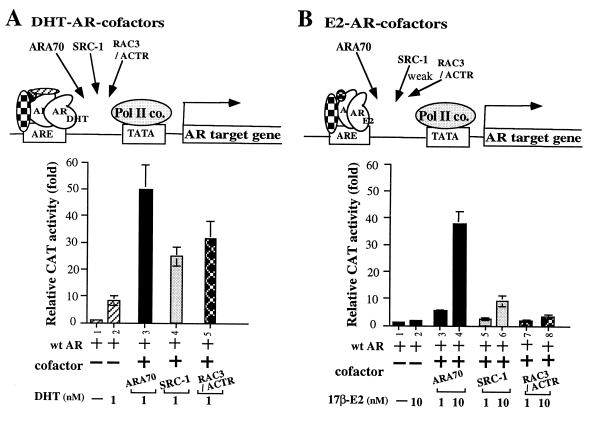
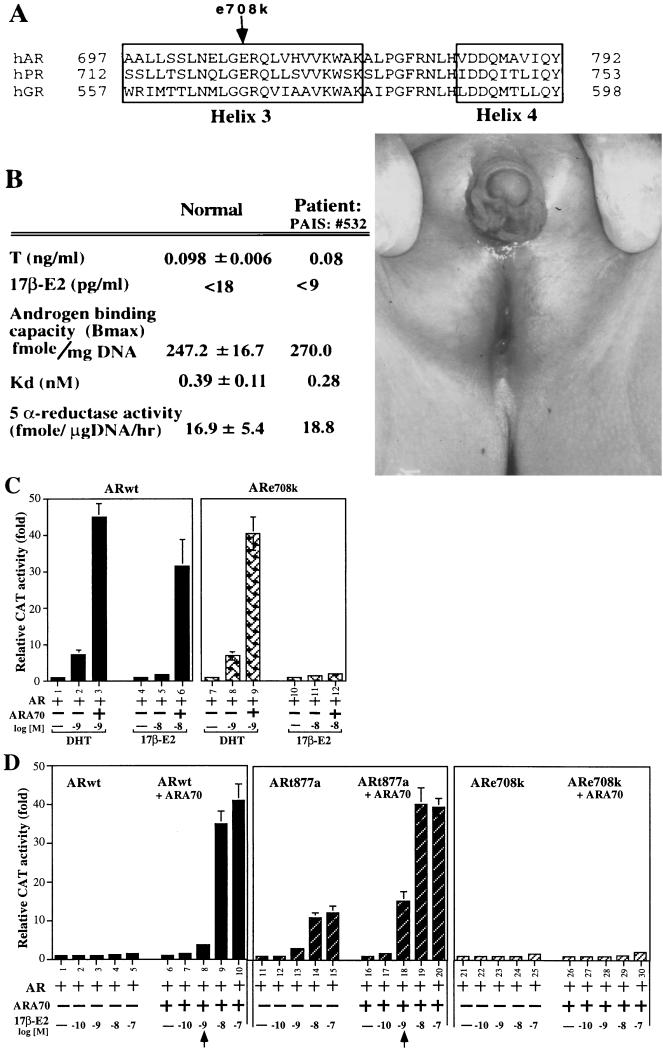
While all three coactivators ARA70, steroid receptor coactivator 1, and RAC3/ACTR can enhance androgen receptor (AR) transcriptional activity at 1 nM dihydrotestosterone, we here demonstrate that only ARA70 can induce AR transcriptional activity >30-fold in the presence of 10 nM 17beta-estradiol (E2), but not diethylstilbestrol. The significance of this newly described E2-induced AR transcriptional activity in DU145 human prostate cancer cells was further strengthened by finding patients with Reifenstein partial-androgen-insensitive syndrome that fail in the E2-AR-ARA70 pathway. Together, our data suggest, for the first time, testosterone/dihydrotestosterone may not be the only ligands for the AR. E2 represents another important natural ligand for AR that may play an essential role for the AR function and the development of the male reproductive system.
Figures
Similar articles
-
Differential induction of androgen receptor transactivation by different androgen receptor coactivators in human prostate cancer DU145 cells.Endocrine. 1999 Oct;11(2):195-202. doi: 10.1385/endo:11:2:195. Endocrine. 1999. PMID: 10709768
-
Interaction of the putative androgen receptor-specific coactivator ARA70/ELE1alpha with multiple steroid receptors and identification of an internally deleted ELE1beta isoform.Mol Endocrinol. 1999 Jan;13(1):117-28. doi: 10.1210/mend.13.1.0214. Mol Endocrinol. 1999. PMID: 9892017
-
Cloning and characterization of a specific coactivator, ARA70, for the androgen receptor in human prostate cells.Proc Natl Acad Sci U S A. 1996 May 28;93(11):5517-21. doi: 10.1073/pnas.93.11.5517. Proc Natl Acad Sci U S A. 1996. PMID: 8643607 Free PMC article.
-
Expression and function of androgen receptor coactivators in prostate cancer.J Steroid Biochem Mol Biol. 2004 Nov;92(4):265-71. doi: 10.1016/j.jsbmb.2004.10.003. Epub 2004 Dec 19. J Steroid Biochem Mol Biol. 2004. PMID: 15663989 Review.
-
Identification and characterization of androgen receptor associated coregulators in prostate cancer cells.J Biol Regul Homeost Agents. 2001 Apr-Jun;15(2):123-9. J Biol Regul Homeost Agents. 2001. PMID: 11501969 Review.
Cited by
-
Expression and function of nuclear receptor co-activator 4: evidence of a potential role independent of co-activator activity.Cell Mol Life Sci. 2012 Dec;69(23):3895-909. doi: 10.1007/s00018-012-1000-y. Epub 2012 May 5. Cell Mol Life Sci. 2012. PMID: 22562579 Free PMC article. Review.
-
Aberrant activation of the androgen receptor by NF-kappaB2/p52 in prostate cancer cells.Cancer Res. 2010 Apr 15;70(8):3309-19. doi: 10.1158/0008-5472.CAN-09-3703. Epub 2010 Apr 13. Cancer Res. 2010. PMID: 20388792 Free PMC article.
-
Synthesis and phosphorylation of androgen receptor of the mouse brain cortex and their regulation by sex steroids during aging.Mol Cell Biochem. 2000 Jan;203(1-2):95-101. doi: 10.1023/a:1007064307220. Mol Cell Biochem. 2000. PMID: 10724337
-
Systems Oncology: Bridging Pancreatic and Castrate Resistant Prostate Cancer.Pathol Oncol Res. 2019 Oct;25(4):1269-1277. doi: 10.1007/s12253-018-0467-8. Epub 2018 Sep 16. Pathol Oncol Res. 2019. PMID: 30220022 Review.
-
A unifying biology of sex steroid-induced apoptosis in prostate and breast cancers.Endocr Relat Cancer. 2018 Feb;25(2):R83-R113. doi: 10.1530/ERC-17-0416. Epub 2017 Nov 21. Endocr Relat Cancer. 2018. PMID: 29162647 Free PMC article. Review.
References
-
- Pratt W B, Welsh M J. Semin Cell Biol. 1994;5:83–93. - PubMed
-
- Tsai M J, O’Malley B W. Annu Rev Biochem. 1994;63:451–486. - PubMed
-
- Jacq X, Brou C, Lutz Y, Davidson I, Chambon P, Tora L. Cell. 1994;79:107–117. - PubMed
-
- Onate S A, Tsai S Y, Tsai M J, O’Malley B W. Science. 1995;270:1354–1357. - PubMed
-
- Horlein A J, Naar A M, Heinzel T, Torchia J, Gloss B, Kurokawa R, Ryan A, Kamei Y, Soderstrom M, Glass C K, et al. Nature (London) 1995;377:397–404. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
