

doi: 10.1073/pnas.95.10.5579.
Angiostatin induces endothelial cell apoptosis and activation of focal adhesion kinase independently of the integrin-binding motif RGD
Affiliations
- PMID: 9576925
- PMCID: PMC20420
- DOI: 10.1073/pnas.95.10.5579
Item in Clipboard
Angiostatin induces endothelial cell apoptosis and activation of focal adhesion kinase independently of the integrin-binding motif RGD
Proc Natl Acad Sci U S A.
.
Abstract
Angiostatin, a fragment of plasminogen, has been identified and characterized as an endogenous inhibitor of neovascularization. We show that angiostatin treatment of endothelial cells in the absence of growth factors results in an increased apoptotic index whereas the proliferation index is unchanged. Angiostatin also inhibits migration and tube formation of endothelial cells. Angiostatin treatment has no effect on growth factor-induced signal transduction but leads to an RGD-independent induction of the kinase activity of focal adhesion kinase, suggesting that the biological effects of angiostatin relate to subversion of adhesion plaque formation in endothelial cells.
Figures
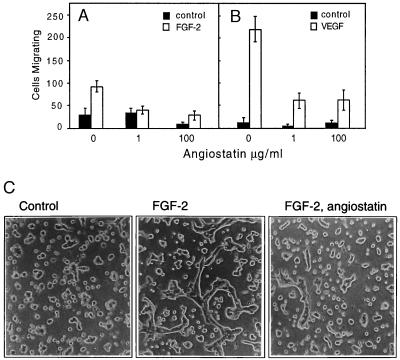
(A and B) Angiostatin attenuates migration of HUVE cells toward FGF-2 (A) and VEGF (B). Cell migration was measured in a Boyden chamber, and the cells migrating through the filter during a 4-h incubation were stained and counted. (C) Angiostatin treatment of murine brain endothelial cells leads to diminished formation of tubes in three-dimensional collagen gels. Murine brain endothelial cells were seeded out on a solidified collagen gel and covered with a second layer of collagen. After 10 h of treatment in the presence or absence of growth factor and angiostatin, the cells were examined for formation of tube-like structures. The cells were treated as indicated with 10% fetal calf serum alone (Left), with 10% fetal calf serum and 5 ng/ml FGF-2 (Center), or pretreated for 72 h with 2.5 μg/ml of angiostatin in Ham’s F-12, 10% fetal bovine serum, followed by inclusion of angiostatin in the collagen gel and stimulation of tube formation by 5 ng/ml FGF-2 (Right). Angiostatin treatment led to 79% decreased formation of tubes, from 305 μm total tube length in the absence (−angiostatin) to 65 μm total tube length in the presence (+) of angiostatin. (×200.)
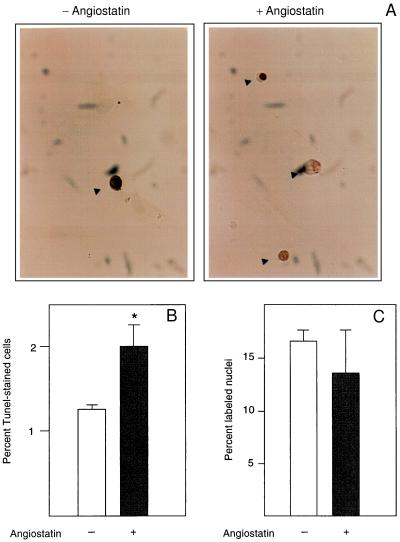
(A) Angiostatin induces apoptosis but has no effect on DNA synthesis in endothelial cells. BCE cells cultured in the presence of 10% serum on coverslips were kept in the absence (−) and presence (+) of 2.5 μg/ml angiostatin for 3 days, at which point the cells were fixed and processed for TUNEL staining (12), to detect cells undergoing programmed cells death. Cells stained positive in the TUNEL assay are indicated by arrowheads. (B) Number of TUNEL-positive cells in the untreated (open bar) and angiostatin-treated (solid bar) cultures. Values are percentage of positive cells ± SEM for four determinations. ∗ indicates P < 0.05. (C) Number of nuclei incorporating [3H]thymidine in the untreated (open bar) and angiostatin-treated (solid bar) samples. Cells were plated on coverslips and cultured in the presence of 10% newborn calf serum with or without 2.5 μg/ml angiostatin for 3 days. During the last 2 h of culture, 1 μCi/ml [3H]thymidine was present in the culture medium. The cells were washed, fixed, and processed for autoradiographic determination of [3H]thymidine incorporation. Values are percentage of labeled nuclei ± SEM for three determinations.

Tyrosine phosphorylation of the adapter protein Shb and activation of mitogen-activated protein (MAP) kinases in response to FGF-2 occur in the presence of angiostatin. BCE cells in DME/10% newborn calf serum were cultured for 24 h with or without 2.5 μg/ml angiostatin, and in some cultures this was followed by 10-min stimulation with 100 ng/ml FGF-2. SDS-denatured cell lysates were immunoprecipitated by using affinity-purified anti-Shb antibodies, followed by SDS/PAGE, transfer to filter, and immunoblotting with antiphosphotyrosine antibodies (PY20, Upper). Immunoblotting of cell extracts separated by SDS/PAGE was performed, followed by transfer to filter and incubation with antibodies specifically recognizing activated MAP kinases Erk 1 and 2 (Lower).
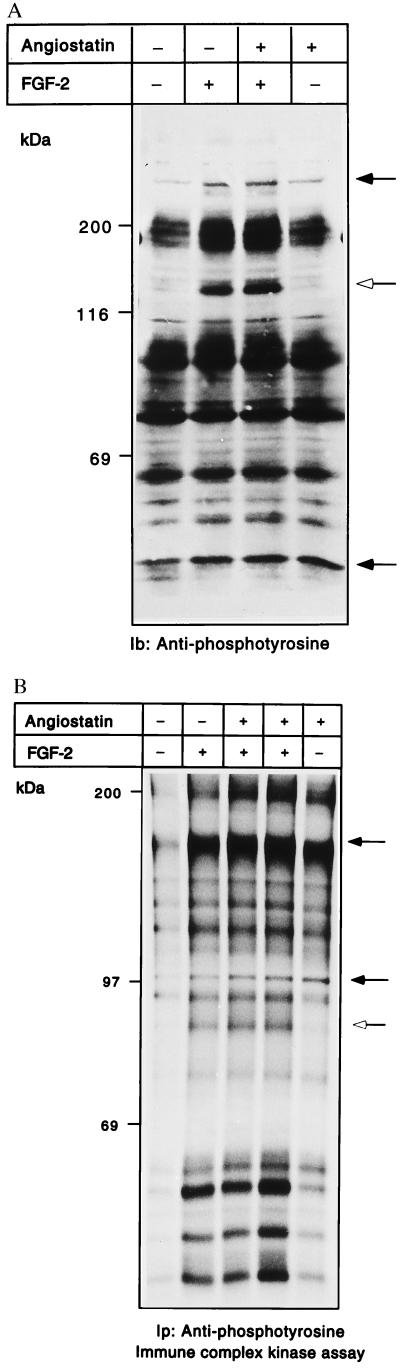
(A and B) Angiostatin treatment leads to induction of kinase activity and tyrosine phosphorylation. BCE cells were cultured in the absence or presence of 2.5 μg/ml angiostatin in the growth medium for 24 h; during the last 10 min, cultures were treated with 100 ng/ml FGF-2 as indicated. Samples of cell extracts were analyzed by immunoblotting by using antiphosphotyrosine antibodies (A). Open arrow indicates a 140-kDa tyrosine-phosphorylated species; solid arrows indicate increased tyrosine phosphorylation in response to angiostatin. PAE cells were similarly treated and cell lysates were used for immunoprecipitation with antiphosphotyrosine antibodies, followed by in vitro kinase assays on the immobilized beads in the presence of [γ-32P]ATP (B). The samples were separated by SDS/PAGE, and the gel was treated with 1 M KOH to reduce phosphorylation on Ser. Solid arrows indicate components with increased 32P-labeled radioactivity in response to angiostatin; open arrow indicates a component that is not affected by angiostatin.
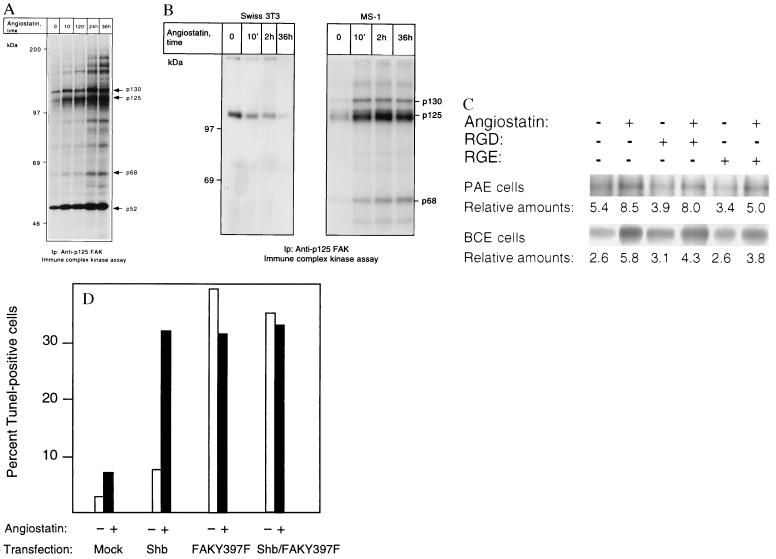
(A and B) p125 FAK is activated in response to angiostatin treatment only in endothelial cells. PAE/FGFR-1 cells (A) and Swiss 3T3 cells and murine pancreas endothelial cells (MS-1; B) were treated as indicated with 2.5 μg/ml of angiostatin for different time periods and processed for immunoprecipitation by using a mAb against p125 FAK. The immobilized immunoprecipitates were subjected to in vitro kinase assays in the presence of [γ-32P]ATP and analyzed by SDS/PAGE. The migration of 130-, 125-, and 68-kDa components that could correspond to Cas, FAK, and paxillin, respectively, as well as other angiostatin-induced phosphorylated components are indicated by arrows, and the relative masses of marker proteins run in parallel are indicated to the left. (C) Treatment with 1 mM RGD-containing peptide fails to block angiostatin-induced FAK activation. PAE/FGFR-1 cells or BCE cells were treated for 10 min with 2.5 μg/ml of angiostatin in the presence and absence of 1 mM peptide containing the RGD motif (Gly-Arg-Gly-Asp-Thr-Pro; Bachem) or with a control peptide containing RGE instead of RGD. The cells were lysed and processed for immunoprecipitation with anti-FAK mAb, followed by in vitro kinase assay. (D) Effects of angiostatin and dominant-negative FAK-Y397F on BCE cell survival. BCE cells were transfected by using the Lipofectamine procedure using lipofectamine alone (mock), Shb cDNA in pcDNA1, and FAK-Y397F in pCMV (a kind gift from J. T. Parsons, University of Virginia) as indicated. The transfected cells were cultured on coverslips in the absence or presence of 2.5 μg/ml angiostatin for 3 days, after which the cells were processed for TUNEL staining and the percentage of stained nuclei were counted.
References
-
- Folkman J. Nat Med. 1995;1:27–31. - PubMed
-
- O’Reilly M S, Holmgren L, Shing Y, Chen C, Rosenthal R A, Moses M, Lane W S, Cao Y, Sage E H, Folkman J. Cell. 1994;79:315–328. - PubMed
-
- O’Reilly M S, Holmgren L, Chen C, Folkman J. Nat Med. 1996;2:689–692. - PubMed
-
- Holmgren L, O’Reilly M S, Folkman J. Nat Med. 1996;1:149–153. - PubMed
-
- Pawson T. Nature (London) 1995;373:573–580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
