

Fas-induced proteolytic activation and intracellular redistribution of the stress-signaling kinase MEKK1
- PMID: 9576928
- PMCID: PMC20423
- DOI: 10.1073/pnas.95.10.5595
Fas-induced proteolytic activation and intracellular redistribution of the stress-signaling kinase MEKK1
Abstract
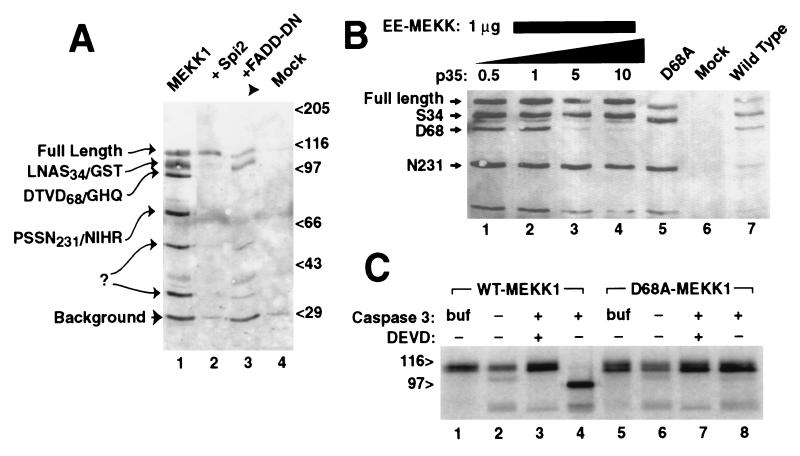
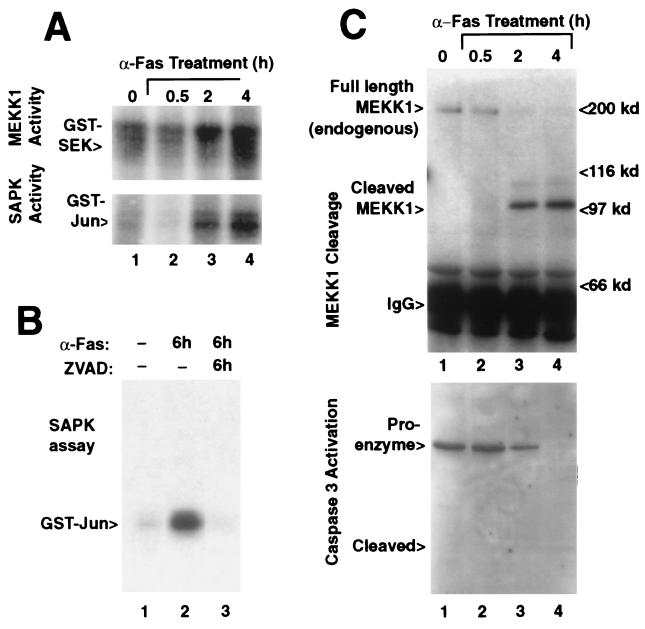
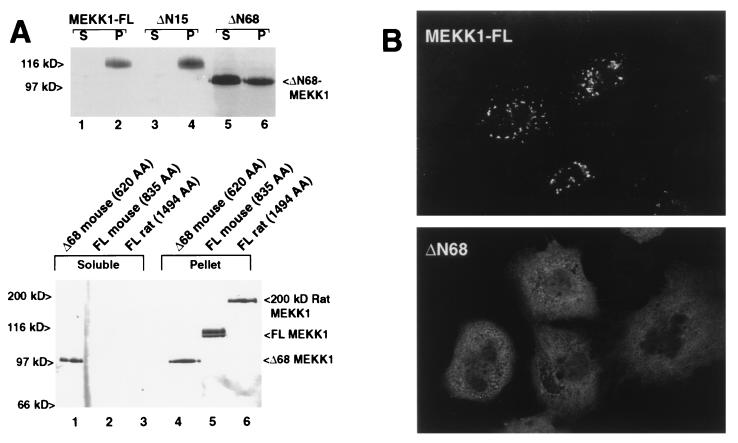
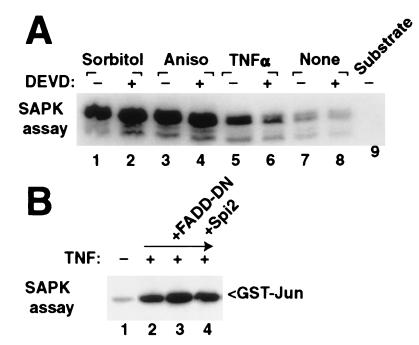
The stress-activated protein kinase (SAPK, alternatively JNK) is activated rapidly by cell stress stimuli such as inflammatory cytokines and oxidative stress, and more slowly by the initiation of the apoptotic cell death response by events such as ligation of the Fas protein. Mitogen-activated protein kinase/Erk kinase kinase-1 (MEKK1) is an activator of SAPK, serving as a SAPK-kinase-kinase through intermediate phosphorylation of the SAPK kinase SEK1. By sequencing proteolytic cleavage products of MEKK1, we found that the proapoptotic protease caspase 3 (CPP32) cleaves MEKK1 after residue D68 both in vivo and in vitro. Cleavage of MEKK1 after D68 is blocked by viral and chemical protease inhibitors. Cleavage of MEKK1 at D68 changes the intracellular distribution of the protein from a Triton-insoluble compartment to a Triton-soluble compartment, reflected in a redistribution from a particulate to a diffuse cytoplasmic staining seen by immunofluorescence. Activation of both SAPK and MEKK1 after Fas ligation is prevented by both viral and chemical caspase 3 inhibitors, which in contrast fail to block activation of SAPK by rapidly acting cell stresses. Stress factor-induced SAPK signaling is not dependent on caspase 3 function. We propose that two mechanisms of stress signaling through MEKK1 exist. One is rapid, independent of proteases, and occurs in the particulate Triton-insoluble compartment. The other is more slowly activated and involves liberation of particulate MEKK1 by proteolytic cleavage and activation by caspase 3.
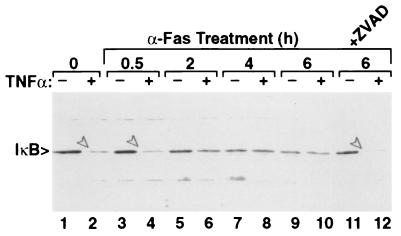
Figures

References
-
- Kyriakis J M, Banerjee P, Nikolakaki E, Dai T, Rubie E A, Ahmad M F, Avruch J, Woodgett J R. Nature (London) 1994;369:156–160. - PubMed
-
- Kyriakis J M, Avruch J. J Biol Chem. 1996;271:24313–24316. - PubMed
-
- Waskiewicz A J, Cooper J A. Curr Opin Cell Biol. 1995;7:798–805. - PubMed
-
- Lange-Carter C A, Pleiman C M, Gardner A M, Blumer K J, Johnson G L. Science. 1993;260:315–319. - PubMed
-
- Sánchez I, Hughes R T, Mayer B, Yee K, Woodgett J R, Avruch J, Kyriakis J M, Zon L I. Nature (London) 1994;372:795–798. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
