

Host cell dependence of viral morphology
- PMID: 9576955
- PMCID: PMC20450
- DOI: 10.1073/pnas.95.10.5746
Host cell dependence of viral morphology
Abstract
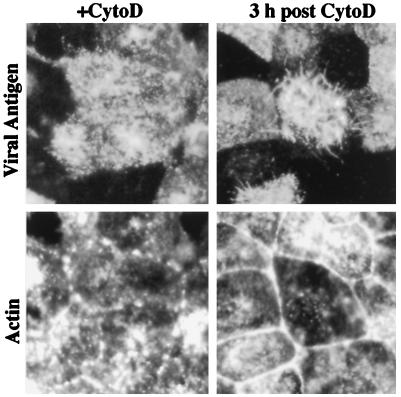
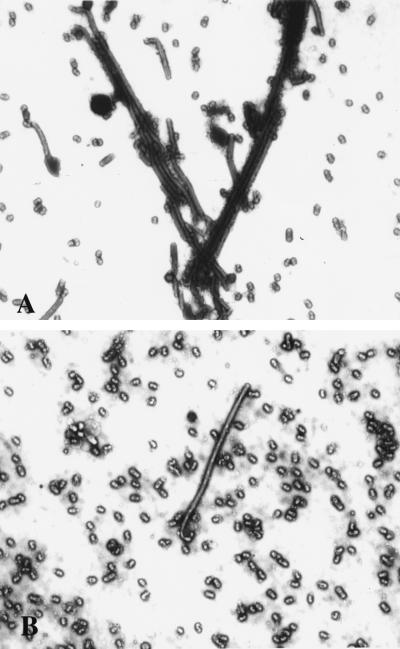
The morphology of influenza virions was found to depend on cellular determinants. Influenza viral filaments up to 30 microm in length were observed to form at high levels on surfaces of various polarized epithelial cell types infected with the A/Udorn/72 virus. In contrast, virions produced by nonpolarized cell types infected with this virus were almost exclusively of spherical morphology. Disruption of the actin microfilament array by cytochalasin D treatment of polarized MDCK cells had a profound effect on viral morphology. Although virus titers and release of spherical particles were not reduced in the presence of cytochalasin D, we observed a 15-fold reduction in the release of filamentous particles. In contrast, the ratio of filaments to spheres produced by infected MDCK cells was not altered by the microtubule-disrupting agent nocodazole. These observations indicate that the polarized cell phenotype and the integrity of the actin microfilament network are important cellular determinants of the morphology of a filamentous influenza virus.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
