

Characterization of interactions between the mammalian polycomb-group proteins Enx1/EZH2 and EED suggests the existence of different mammalian polycomb-group protein complexes
- PMID: 9584199
- PMCID: PMC108940
- DOI: 10.1128/MCB.18.6.3586
Characterization of interactions between the mammalian polycomb-group proteins Enx1/EZH2 and EED suggests the existence of different mammalian polycomb-group protein complexes
Abstract
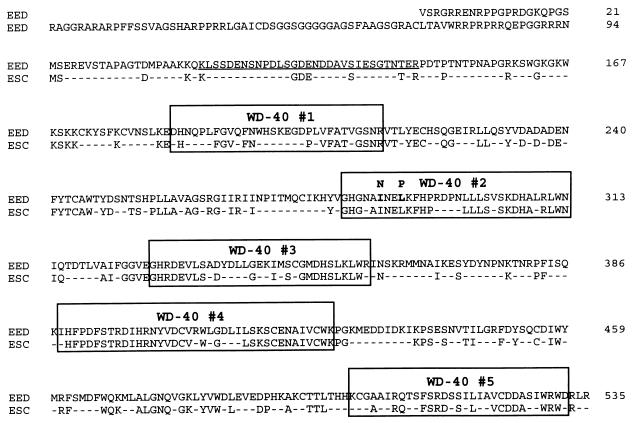
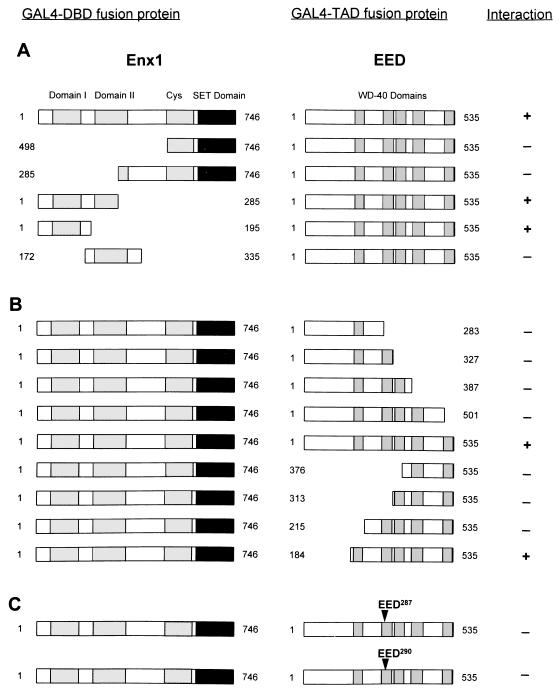
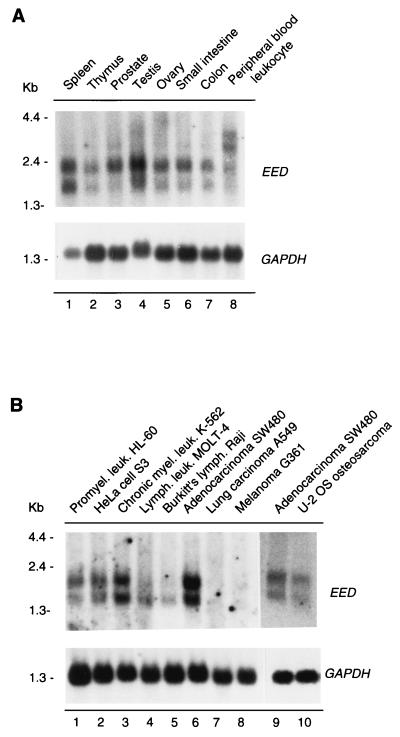
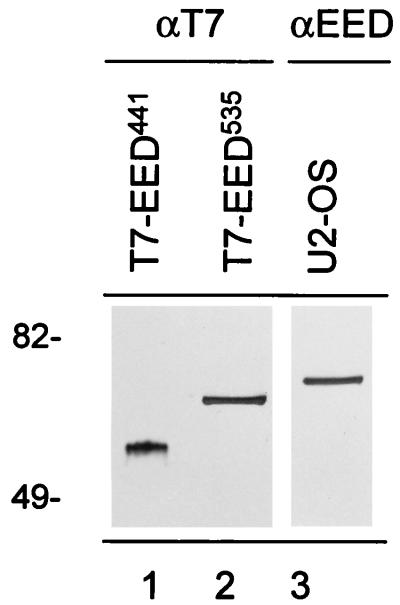
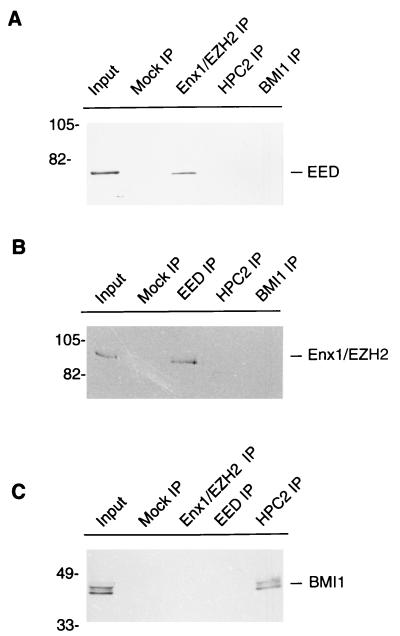
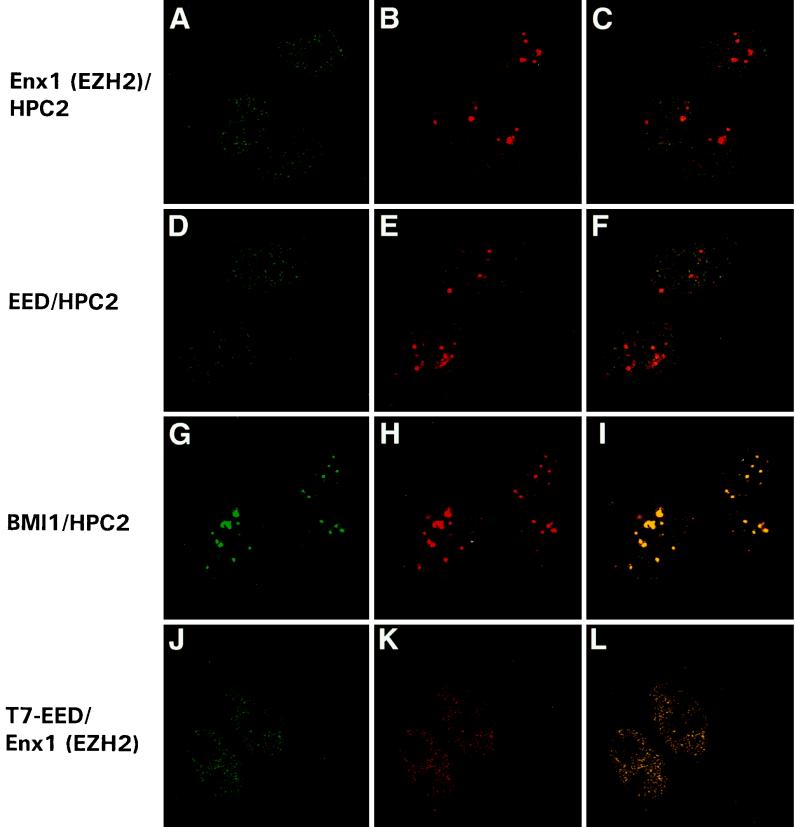
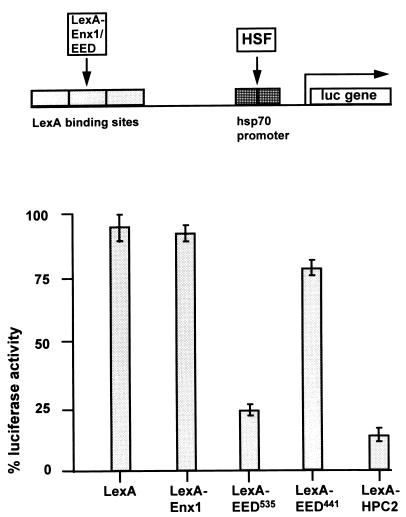
In Drosophila melanogaster, the Polycomb-group (PcG) and trithorax-group (trxG) genes have been identified as repressors and activators, respectively, of gene expression. Both groups of genes are required for the stable transmission of gene expression patterns to progeny cells throughout development. Several lines of evidence suggest a functional interaction between the PcG and trxG proteins. For example, genetic evidence indicates that the enhancer of zeste [E(z)] gene can be considered both a PcG and a trxG gene. To better understand the molecular interactions in which the E(z) protein is involved, we performed a two-hybrid screen with Enx1/EZH2, a mammalian homolog of E(z), as the target. We report the identification of the human EED protein, which interacts with Enx1/EZH2. EED is the human homolog of eed, a murine PcG gene which has extensive homology with the Drosophila PcG gene extra sex combs (esc). Enx1/EZH2 and EED coimmunoprecipitate, indicating that they also interact in vivo. However, Enx1/EZH2 and EED do not coimmunoprecipitate with other human PcG proteins, such as HPC2 and BMI1. Furthermore, unlike HPC2 and BMI1, which colocalize in nuclear domains of U-2 OS osteosarcoma cells, Enx1/EZH2 and EED do not colocalize with HPC2 or BMI1. Our findings indicate that Enx1/EZH2 and EED are members of a class of PcG proteins that is distinct from previously described human PcG proteins.
Figures

References
-
- Alkema M J, Wiegant J, Raap A K, Berns A, van Lohuizen M. Characterization and chromosomal localization of the human proto-oncogene BMI-1. Hum Mol Genet. 1993;2:1597–1603. - PubMed
-
- Alkema M J, Bronk M, Verhoeven E, Otte A P, van’t Veer L J, Berns A, van Lohuizen M. Identification of Bmi1-interacting proteins as constituents of a multimeric mammalian Polycomb complex. Genes Dev. 1997;11:226–240. - PubMed
-
- Carrington E A, Jones R S. The Drosophila Enhancer of zeste gene encodes a chromosomal protein: examination of wild-type and mutant protein distribution. Development. 1996;122:4073–4083. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases