

Regulation of the fission yeast transcription factor Pap1 by oxidative stress: requirement for the nuclear export factor Crm1 (Exportin) and the stress-activated MAP kinase Sty1/Spc1
- PMID: 9585505
- PMCID: PMC316839
- DOI: 10.1101/gad.12.10.1453
Regulation of the fission yeast transcription factor Pap1 by oxidative stress: requirement for the nuclear export factor Crm1 (Exportin) and the stress-activated MAP kinase Sty1/Spc1
Erratum in
- Genes Dev 1998 Aug 15;12(16):2650
Abstract
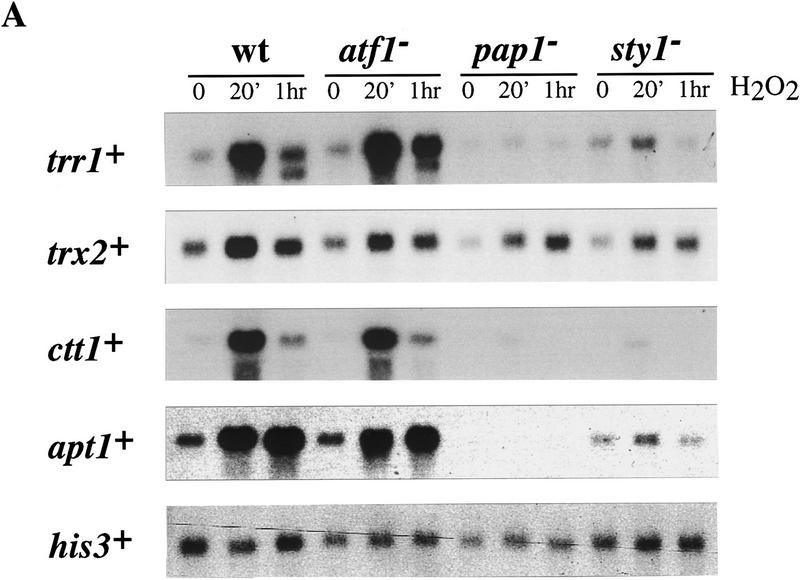
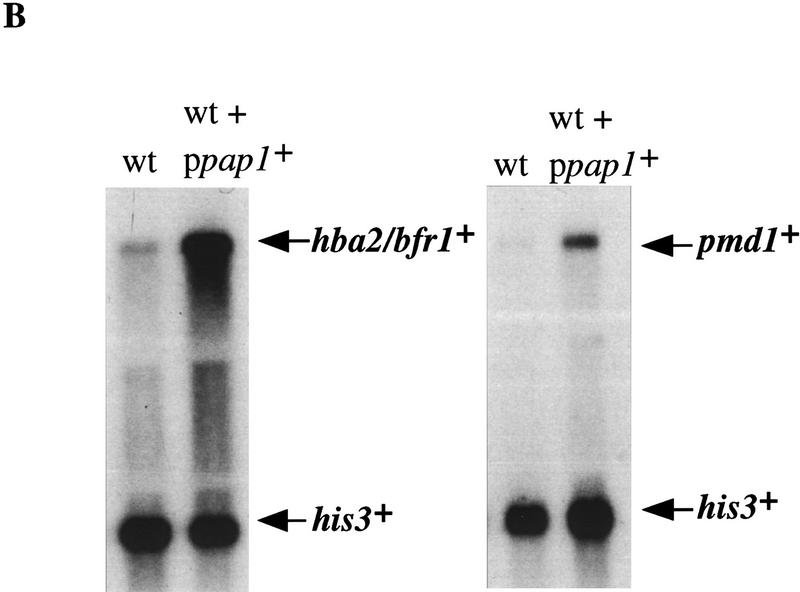
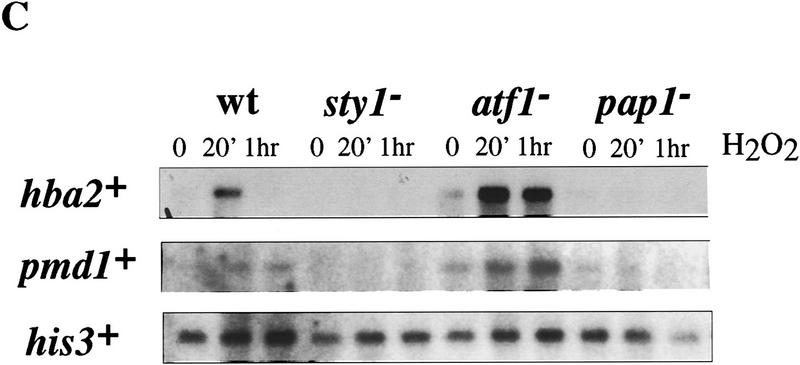
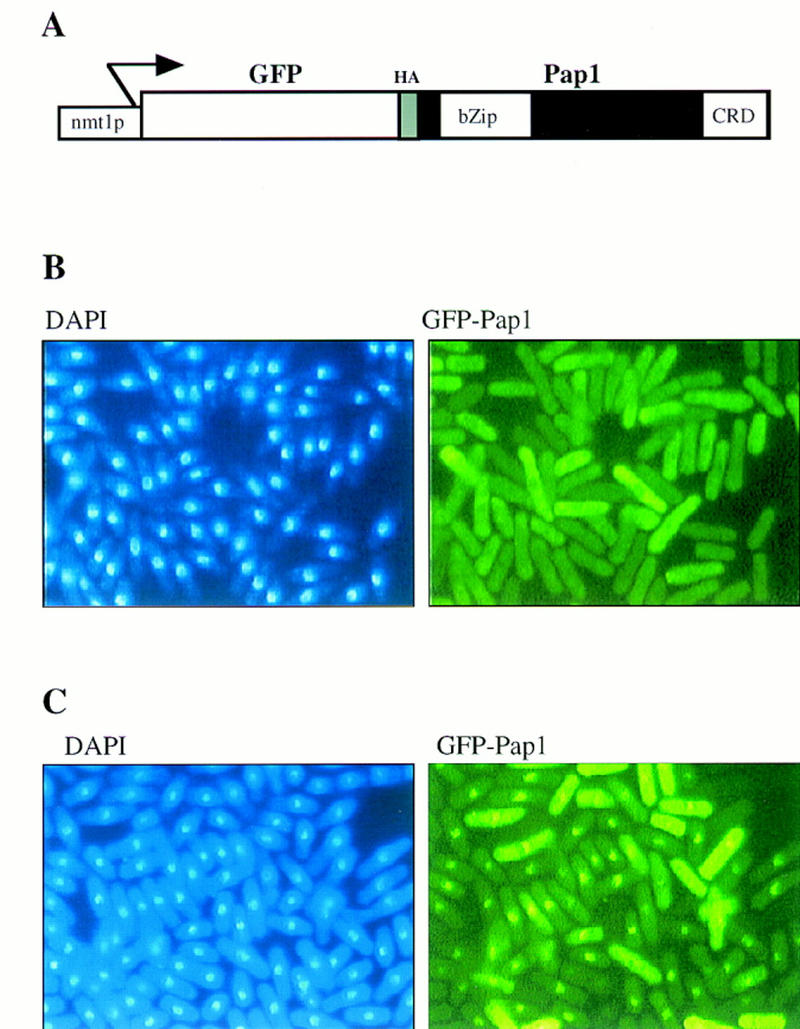
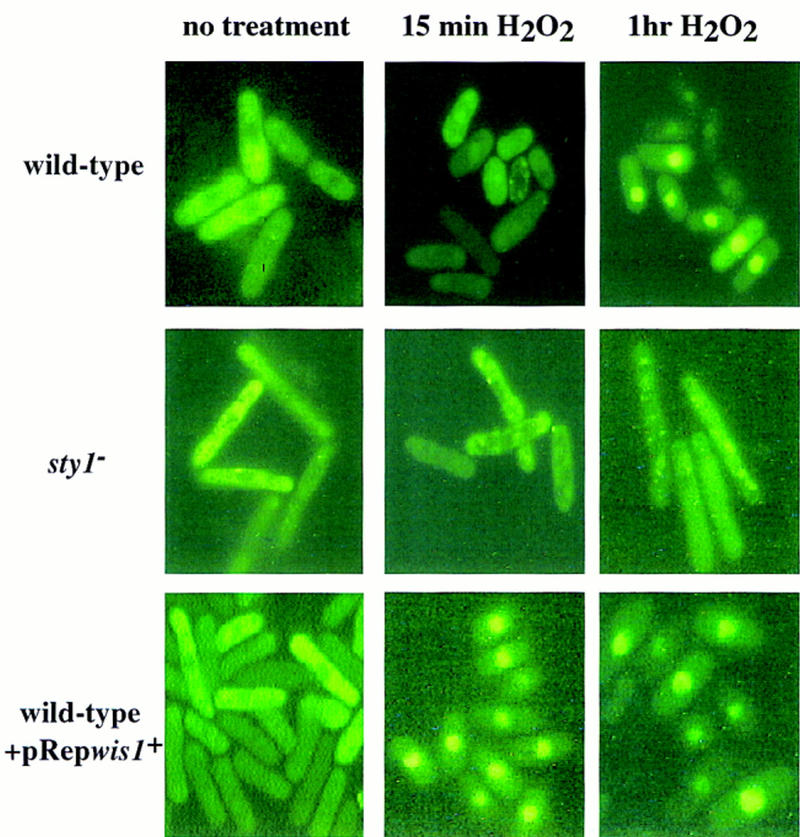
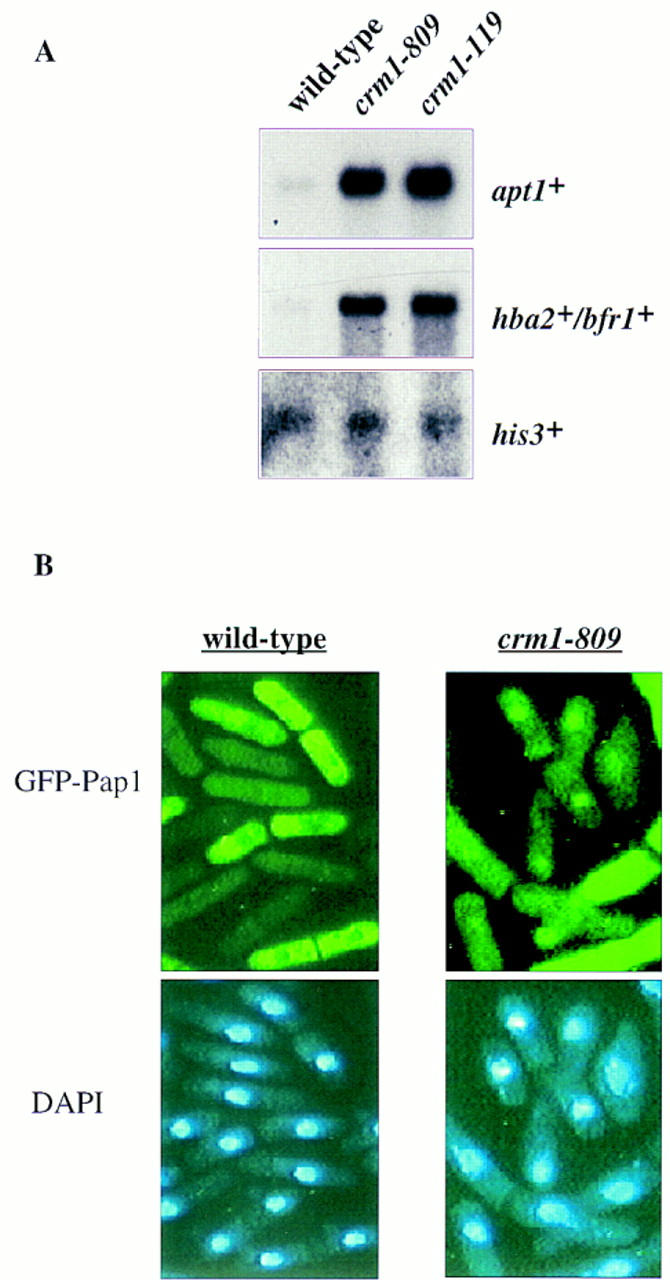
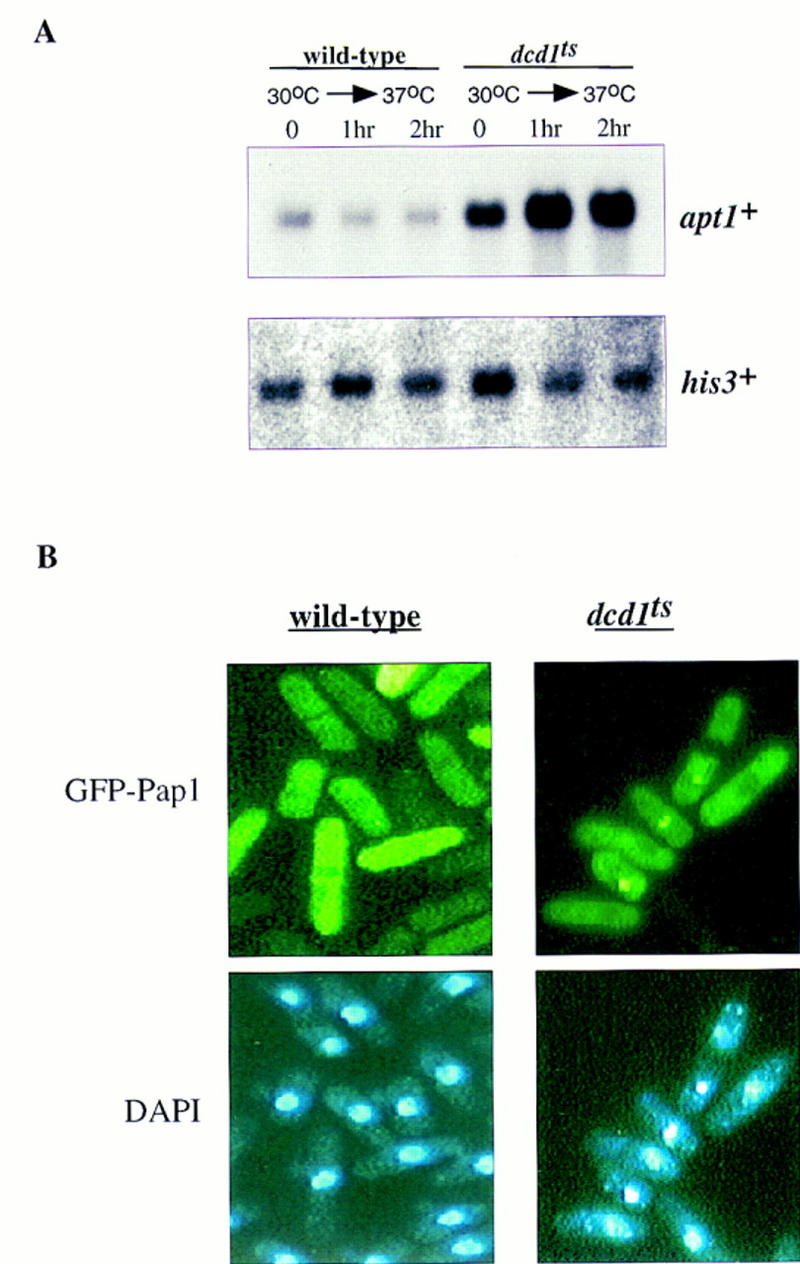
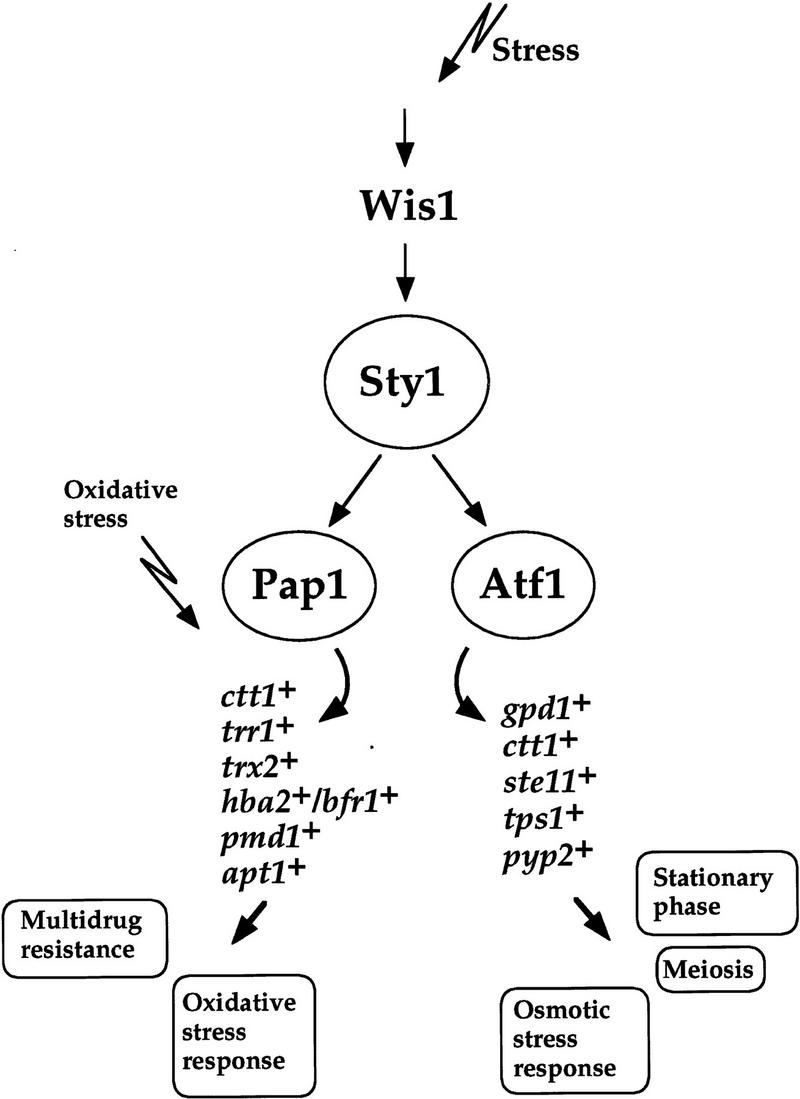
The fission yeast Sty1 stress-activated MAP kinase is crucial for the cellular response to a variety of stress conditions. Accordingly, sty1- cells are defective in their response to nutrient limitation, lose viability in stationary phase, and are hypersensitive to osmotic stress, oxidative stress, and UV treatment. Some of these phenotypes are caused by Sty1-dependent regulation of the Atf1 transcription factor, which controls both meiosis-specific and osmotic stress-responsive genes. However, in this report we demonstrate that the cellular response to oxidative stress and to treatment with a variety of cytotoxic agents is the result of Sty1 regulation of the Pap1 transcription factor, a bZip protein with structural and DNA binding similarities to the mammalian c-Jun protein. We show that both Sty1 and Pap1 are required for the expression of a number of genes involved in the oxidative stress response and for the expression of two genes, hba2+/bfr1+ and pmd1+, which encode energy-dependent transport proteins involved in multidrug resistance. Furthermore, we demonstrate that Pap1 is regulated by stress-dependent changes in subcellular localization. On imposition of oxidative stress, the Pap1 protein relocalizes from the cytoplasm to the nucleus in a process that is dependent on the Sty1 kinase. This relocalization is the result of regulated protein export, rather than import, and involves the Crm1 (exportin) nuclear export factor and the dcd1+/pim1+ gene that encodes an Ran nucleotide exchange factor.
Figures

Comment in
-
SAPKs and transcription factors do the nucleocytoplasmic tango.Genes Dev. 1998 May 15;12(10):1391-7. doi: 10.1101/gad.12.10.1391. Genes Dev. 1998. PMID: 9585499 Review. No abstract available.
References
-
- Alfa C, Fantes P, Hyams J, McLeod M, Warbrick E. Experiments with fission yeast. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1993.
-
- Baeuerle PA, Henkel T. Function and activation of NFκB in the immune system. Annu Rev Immunol. 1994;12:540–546. - PubMed
-
- Brewster JL, de Valoir T, Dwyer ND, Winter E, Gustin MC. An osmosensing signal transduction pathway in yeast. Science. 1993;259:1760–1763. - PubMed
-
- Casso D, Beach D. A mutation in a thioredoxin reductase homolog suppresses p53-induced growth-inhibition in the fission yeast Schizosaccharomyces pombe. Mol & Gen Genet. 1996;252:518–529. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous