

Review
doi: 10.1016/s0167-7799(97)01167-0.
Translating old drugs into new treatments: ribosomal frameshifting as a target for antiviral agents
Affiliations
- PMID: 9586242
- PMCID: PMC7127214
- DOI: 10.1016/s0167-7799(97)01167-0
Item in Clipboard
Review
Translating old drugs into new treatments: ribosomal frameshifting as a target for antiviral agents
Trends Biotechnol.
1998 Apr.
Abstract
Programmed ribosomal frameshifting is used by many viruses to regulate the production of structural and enzymatic proteins. Altering the frameshifting efficiencies disrupts the virus life cycle and eliminates or reduces virus production. Ribosomal frameshifting therefore provides a unique target on which antiviral agents can function. This article describes a series of rapid assay strategies that have been developed and used to identify potential antiviral agents that target programmed -1 ribosomal frameshifting.
Figures
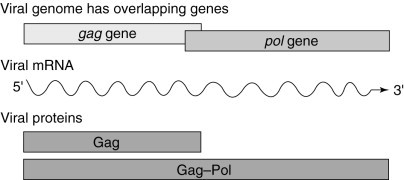
The genomes of viruses that use programmed ribosomal frameshifting are generally organized so that the open reading frames (ORFs) encoding structural proteins (e.g. gag) are 5′ of, and translated prior to, the ORFs encoding proteins with enzymatic functions (e.g. pol). Furthermore, pol is out of frame with respect to gag. The majority (90–98%) of host ribosomes translating the single viral mRNA terminate translation at the gag stop codon, resulting in the synthesis of Gag protein. A minority of ribosomes (2–10%) are induced to shift reading frame at the viral frameshift signal, resulting in the synthesis of the Gag–Pol fusion protein.
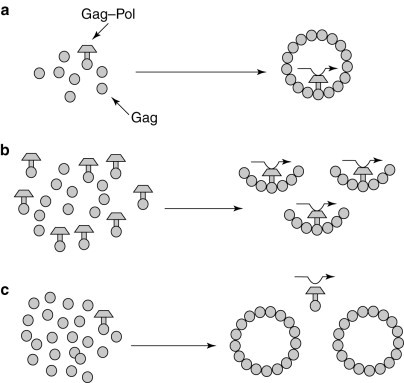
The efficiency of the frameshift determines the ratio of Gag to Gag–Pol proteins available for viral particle morphogenesis. (a) The normal frameshifting efficiency provides the correct ratio of Gag to Gag–Pol. (b) Increased frameshifting leads to the formation of incomplete viral particles. (c) Decreased frameshifting efficiency leads to the formation of Gag–Pol-deficient viral particles that cannot package the viral (+) strand.
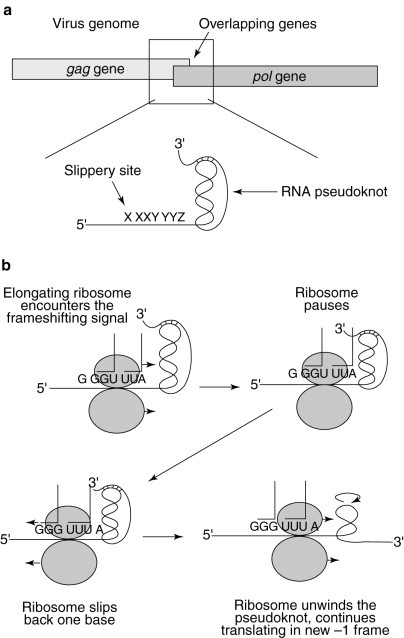
The simultaneous-slippage model of programmed ribosomal −1 frameshifting. (a) The programmed −1 ribosomal frameshift signal is located in the region where the gag and pol ORFs overlap. (b) An elongating ribosome encounters the RNA pseudoknot and pauses over the heptameric slippery site. The tRNAs in the ribosomal A and P sites unpair from the 0-frame codons and slip back one base so that their non-wobble bases rebind with the −1-frame codons. The ribosome unwinds the RNA pseudoknot and continues to translate the mRNA in the new −1 reading frame.
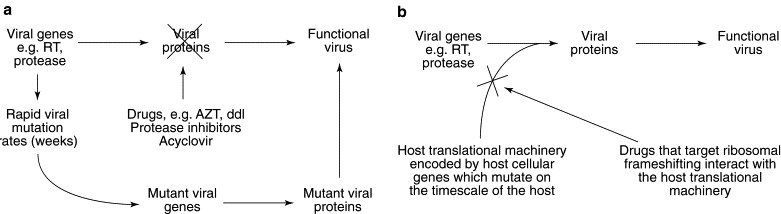
Advantages in targeting programmed −1 ribosomal frameshifting for antiviral therapies. (a) Conventional antiviral drugs target virus-encoded proteins. The rapid viral mutation rates ensure the selection of mutant viral genes encoding viral proteins that can bypass the actions of the drugs, resulting in drug-resistant functional virus. (b) Drugs that target programmed ribosomal frameshifting affect the host translational machinery, which is independent of the viral mutational capacity.
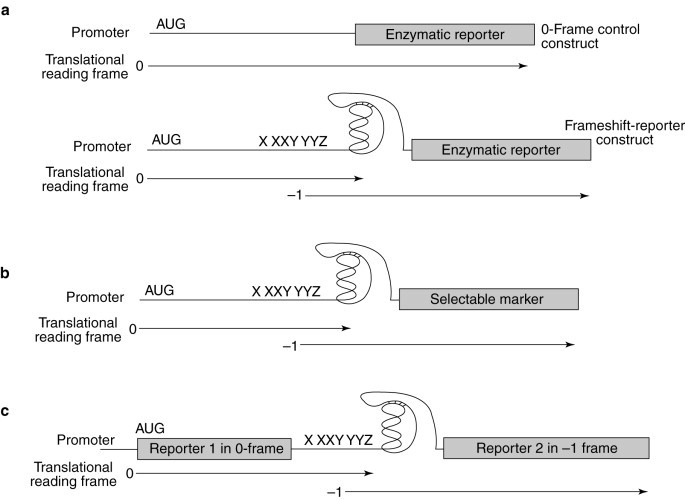
Programmed −1 ribosomal frameshift assay strategies. (a) General strategy for directly measuring the efficiency of programmed −1 ribosomal frameshifting. Pictured are the 0-frame control reporter and −1 ribosomal frameshift constructs. Both systems utilize a reporter protein (the lacZ-encoded β-galactosidase, in vivo, or luciferase, in vitro). In the −1 ribosomal frameshift construct, the reporter is cloned downstream of programmed −1 ribosomal frameshift signals and is in the −1 frame with a translational start site. Synthesis of the reporter protein thus requires a programmed ribosomal frameshift event. The 0-frame controls have the reporter genes cloned in frame with the start site and lack the intervening viral frameshift signals. The programmed ribosomal frameshift efficiency is determined by dividing the level of reporter-protein activity produced from the −1 reporters by that produced from the 0-frame control and multiplying by 100%. Translation of the reporters can occur in intact yeast cells or in translationally competent cell extracts. The effects of a candidate compound on both overall translation and on programmed −1 ribosomal frameshifting can be monitored. (b) General strategy for assays in intact yeast cells that detect changes in programmed −1 ribosomal frameshift efficiencies by using a selectable marker. A gene encoding a selectable marker is cloned downstream of, and in the −1 frame with respect to, a translational start site, such that synthesis of the gene product requires a programmed ribosomal frameshift event. Changes in programmed −1 ribosomal frameshift efficiencies are detected by monitoring the ability of cells to grow (positive selection) or not (negative selection) in the presence of a candidate compound. (c) Bicistronic constructs for measuring changes in programmed −1 ribosomal frameshifting efficiencies. Reporter-gene 1 constitutes the 0-frame control and can be used to monitor the effect of a compound on overall translation. Translation of reporter-gene 2 requires a programmed ribosomal frameshift event. Changes in programmed −1 ribosomal frameshift efficiencies are detected by monitoring for changes in the ratios between the activities of the two reporter proteins.
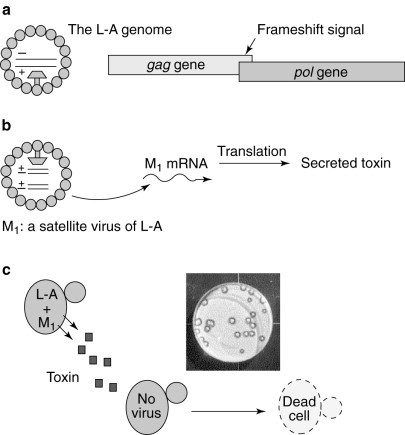
The yeast `killer-virus' system: an ideal assay system. The killer-virus system is composed of the L-A dsRNA helper virus and the M1 dsRNA satellite virus. (a) The gag and pol ORFs overlap in the L-A genome, and a programmed −1 ribosomal frameshift is used to synthesize the Gag and Gag–Pol fusion proteins to produce a functional viral particle containing the L-A dsRNA. (b) The dsRNA genome of the M1 satellite virus is encapsidated inside L-A-encoded viral particles; the M1 mRNA encodes a secreted toxin. (c) Cells harbouring both L-A and M1 secrete the toxin and are immune to its action, whereas virus-free cells are sensitive to the toxin. A picture of the yeast-killer assay is shown, in which cells harbouring L-A and M1 were replica-plated onto a lawn of virus-free cells. A ring of growth inhibition is indicative of the killer activity of the cells harbouring both L-A and M1.
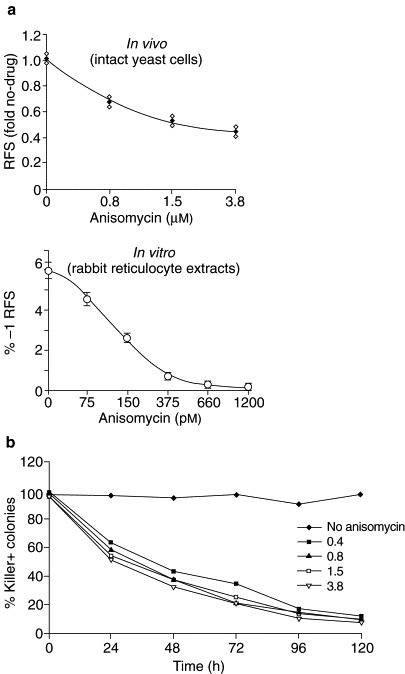
Anisomycin specifically decreases −1 ribosomal frameshifting efficiency and has antiviral activity. (a) The effects of anisomycin on programmed ribosomal frameshifting were assayed in intact yeast cells in vivo, and in translationally competent rabbit reticulocyte extracts in vitro. The indicated drug concentrations were subinhibitory for cell growth and division (in vivo), and for overall translation (both in vivo and in vitro) (not shown). Anisomycin specifically inhibits programmed −1 ribosomal frameshifting in both systems. (b) Yeast cells harbouring the L-A and M1 viruses were cultured in the presence of the indicated concentrations of anisomycin for the indicated times, after which the percentage of cells retaining killer activity was determined.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
