

Induction of a specific olfactory memory leads to a long-lasting activation of protein kinase C in the antennal lobe of the honeybee
- PMID: 9592115
- PMCID: PMC6792810
- DOI: 10.1523/JNEUROSCI.18-11-04384.1998
Induction of a specific olfactory memory leads to a long-lasting activation of protein kinase C in the antennal lobe of the honeybee
Abstract
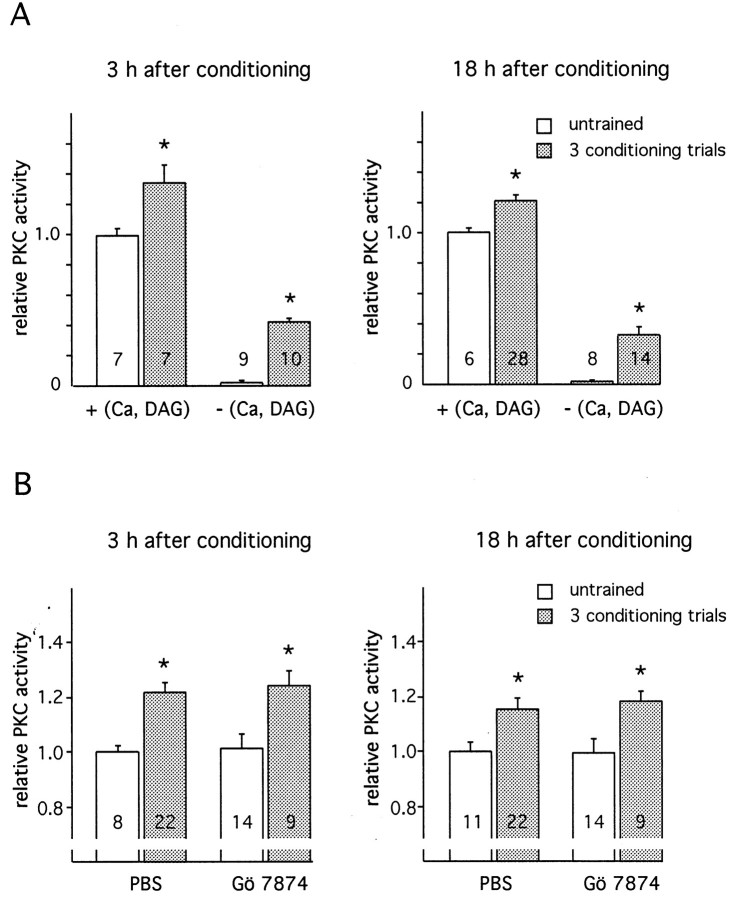
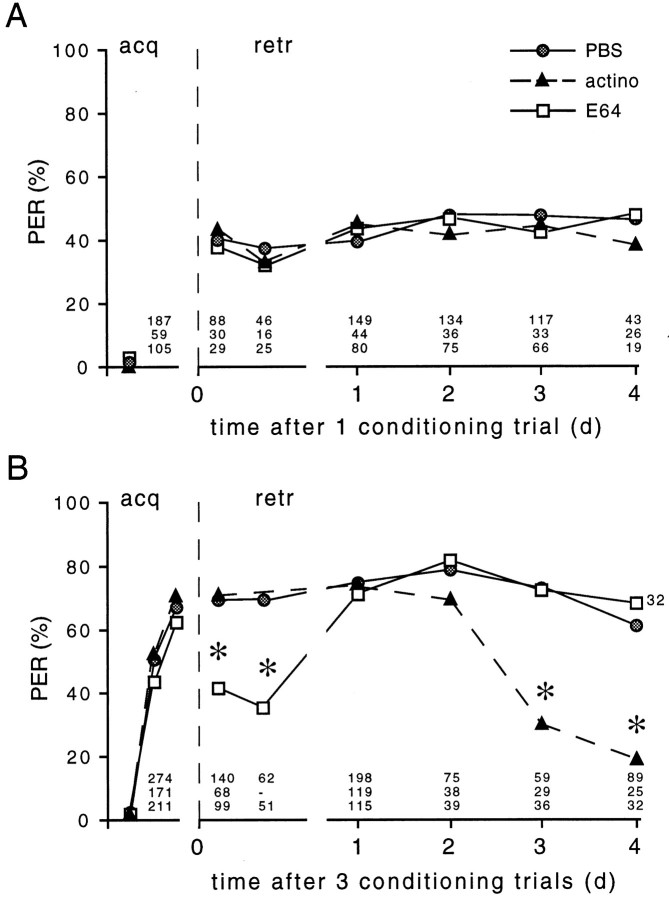
In this study we investigated the role of protein kinase C (PKC) in associative learning of Apis mellifera. Changes in PKC activity induced by olfactory conditioning were measured in the antennal lobes, a brain structure involved in associative learning. Multiple conditioning trials inducing a memory different from that induced by a single conditioning trial specifically cause an increase in PKC activity. This increase begins 1 hr after conditioning, lasts up to 3 d, and is attributable to an increased level of constitutive PKC. The increased level of constitutive PKC consists of an early proteolysis-dependent phase and a late phase that requires RNA and protein synthesis. Inhibition of the pathways resulting in constitutive PKC selectively impairs distinct phases of multiple-trial induced memory. The inhibition of the proteolytic mechanism has an instant effect on an early phase of multiple-trial induced memory but does not affect acquisition and the late phase of memory. Blocking of the transient PKC activation during conditioning does not affect the induction of memory formation. Thus, the constitutive PKC in the antennal lobe seems to contribute to the early phase of memory that is induced by multiple-trial conditioning.
Figures




References
-
- Abel T, Nguyen PV, Barad M, Deuel AS, Kandel E, Bourtchouladze R. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell. 1997;88:615–626. - PubMed
-
- Akers RF, Lovinger DM, Colley PA, Linden DJ, Routtenberg A. Translocation of protein kinase C activity may mediate hippocampal long-term potentiation. Science. 1986;231:587–589. - PubMed
-
- Altfelder K, Müller U, Menzel R. Ca2+/calmodulin and Ca2+/phospholipid-dependent protein kinases in the neuronal tissue of the honeybee Apis Mellifera. Insect Biochem. 1991;21:479–486.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical