

In vivo binding of immunoglobulin M to the surfaces of Babesia bigemina-infected erythrocytes
- PMID: 9596768
- PMCID: PMC108290
- DOI: 10.1128/IAI.66.6.2922-2927.1998
In vivo binding of immunoglobulin M to the surfaces of Babesia bigemina-infected erythrocytes
Abstract
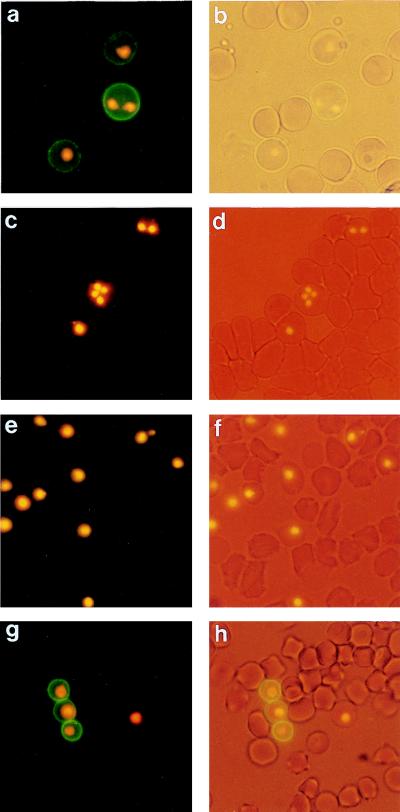
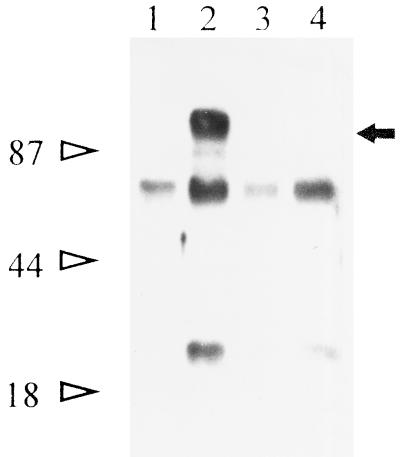
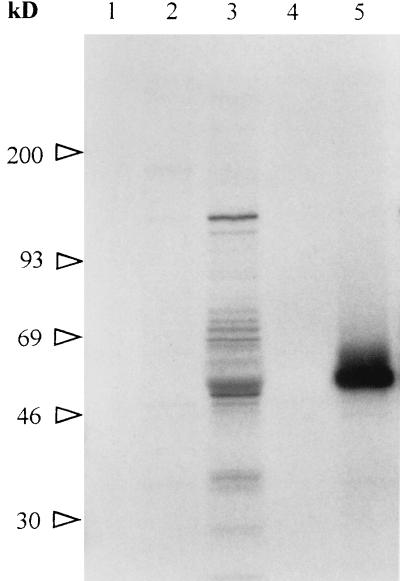
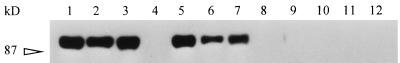
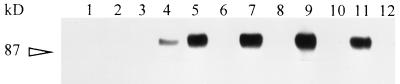
Babesia bigemina infection of mature bovine erythrocytes results in new proteins specifically exposed on the parasitized cell surface. Monoclonal antibody (MAb) 64/32 binds a protein, designated p94, on B. bigemina-infected erythrocytes but not on either uninfected or B. bovis-parasitized erythrocytes. However, p94 was not encoded by B. bigemina and was not a parasite-modified erythrocyte membrane protein. In contrast, we showed that p94 could be eluted from the infected erythrocyte surface and was identified as specifically bound immunoglobulin M (IgM) heavy chain for the following reasons: (i) MAb 64/32 bound a reduced molecule of 94 kDa in both infected erythrocyte lysates and normal bovine serum; (ii) MAb 64/32 bound a 94-kDa molecule in reduced preparations of purified IgM; (iii) an anti-bovine mu heavy-chain MAb, BIg73, reacted specifically with the surface of infected erythrocytes and bound the 94-kDa molecule in lysates of infected erythrocytes, normal bovine serum, and purified IgM; and (iv) immunoprecipitation of infected erythrocyte lysates with MAb 64/32 depleted the 94-kDa antigen bound by anti-mu MAb BIg73 and vice versa. Binding of IgM to the infected erythrocyte surface was detected in vivo early in acute parasitemia and occurred during both the trophozoite and merozoite stages of intraerythrocytic parasitism. The common feature of IgM binding to the parasitized erythrocyte surface among otherwise genetically and antigenically distinct B. bigemina strains is suggestive of an advantageous role in parasite survival in vivo.
Figures

References
-
- Adams J H, Hudson D E, Torii M, Ward G E, Wellems T E, Aikawa M, Miller L H. The Duffy receptor family of Plasmodium knowlesi is located within the micronemes of invasive malaria merozoites. Cell. 1990;63:141–153. - PubMed
-
- Aikawa M, Rabbege J, Uni S, Ristic M, Miller L H. Structural alteration of the membrane of the erythrocytes infected with Babesia bovis. Am J Trop Med Hyg. 1985;34:45–49. - PubMed
-
- Baruch D, Pasloske B L, Singh H B, Bi X, Ma X C, Feldman M, Taraschi T F, Howard R J. Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 1995;82:77–87. - PubMed
-
- Berendt A R, Ferguson D J P, Gardner J, Turner G, Rowe A, McCormick C, Roberts D, Craig A, Pinches R, Elford B C, Newbold C I. Molecular mechanisms of sequestration in malaria. Parasitol. 1990;108:S19–S28. - PubMed
-
- Crandall I, Guthrie N, Sherman I W. Plasmodium falciparum: sera of individuals living in a malaria-endemic region recognize peptide motifs of the human erythrocyte anion transport protein. Am J Trop Med Hyg. 1995;52:450–452. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
