

Trypanosoma cruzi infection in tumor necrosis factor receptor p55-deficient mice
- PMID: 9596773
- PMCID: PMC108295
- DOI: 10.1128/IAI.66.6.2960-2968.1998
Trypanosoma cruzi infection in tumor necrosis factor receptor p55-deficient mice
Abstract
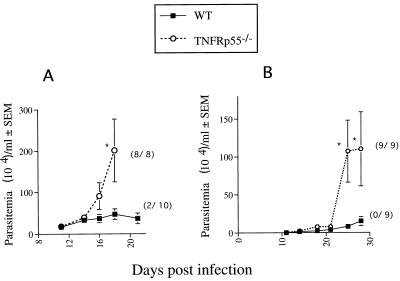
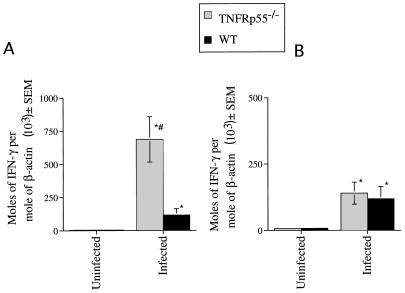
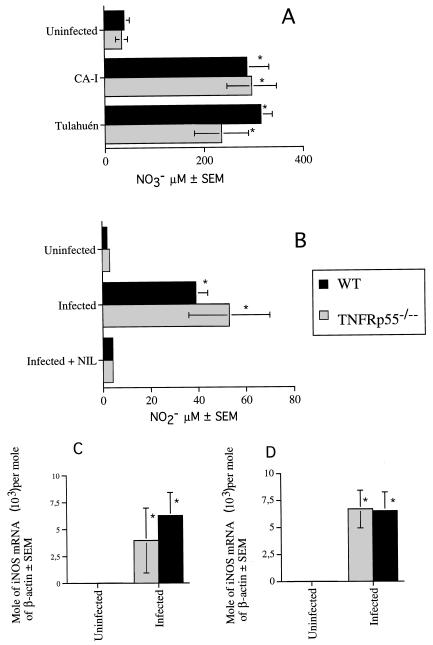
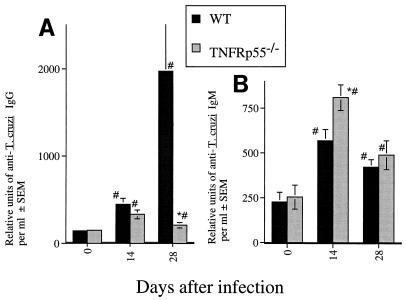
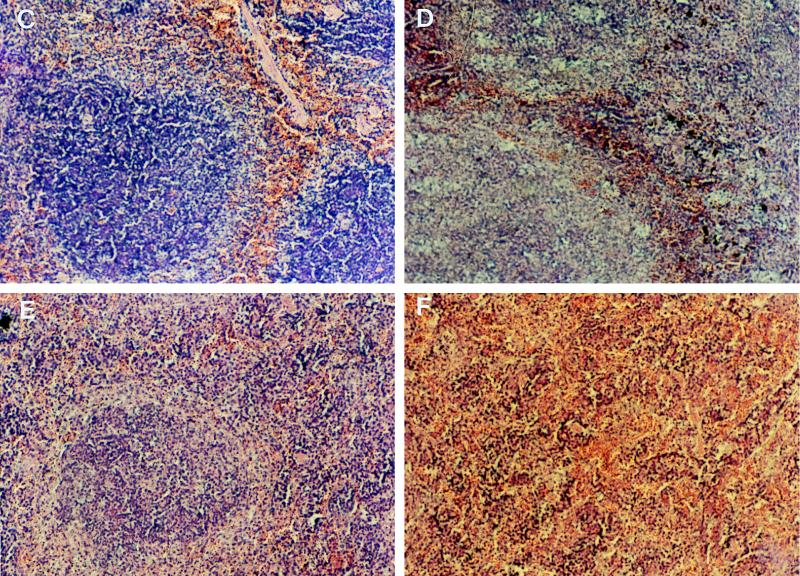
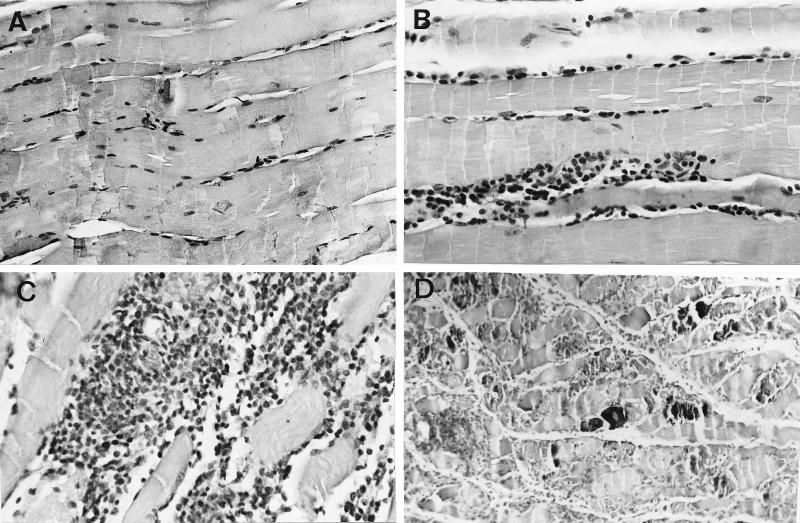
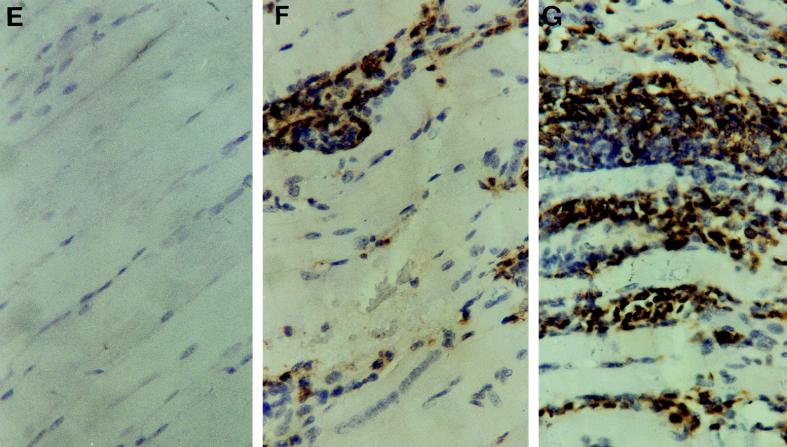
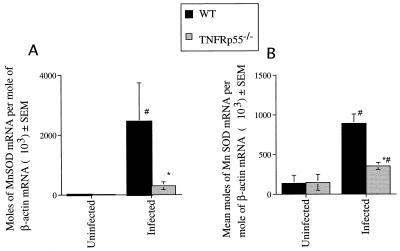
Tumor necrosis factor receptor p55 (TNFRp55) mediates host resistance to several pathogens by allowing microbicidal activities of phagocytes. In the studies reported here, TNFRp55-/- mice infected with the intracellular parasite Trypanosoma cruzi showed clearly higher parasitemia and cumulative mortality than wild-type (WT) controls did. However, gamma interferon (IFN-gamma)-activated macrophages from TNFRp55-/- mice produced control levels of nitric oxide and killed the parasite efficiently in vitro. Trypanocidal mechanisms of nonphagocytic cells (myocardial fibroblasts) from both TNFRp55-/- and WT mice were also activated by IFN-gamma in a dose-dependent way. However, IFN-gamma-activated TNFRp55-/- nonphagocytes showed less effective killing of T. cruzi than WT control nonphagocytes, even when interleukin 1beta (IL-1beta) was added as a costimulator. In vivo, T. cruzi-infected TNFRp55-/- mice and WT mice released similar levels of NO and showed similar levels of IFN-gamma mRNA and inducible nitric oxide synthase mRNA in their tissues. Instead, increased susceptibility to T. cruzi of TNFRp55-/- mice was associated with reduced levels of parasite-specific immunoglobulin G (IgG) (but not IgM) antibodies during infection, which is probably linked to abnormal B-cell differentiation in secondary lymphoid tissues of the mutant mice. Surprisingly, T. cruzi-infected TNFRp55-/- mice showed increased inflammatory and necrotic lesions in several tissues, especially in skeletal muscles, indicating that TNFRp55 plays an important role in controlling the inflammatory process. Accordingly, levels of Mn2+ superoxide dismutase mRNA, a TNF-induced enzyme which protects the cell from the toxic effects of superoxide, were lower in mutant than in WT infected mice.
Figures
References
-
- Bancroft G J, Sheehan K C F, Shreiber R D, Unanue E R. Tumour necrosis factor is involved in the T-cell independent pathway of macrophage activation in scid mice. J Immunol. 1989;143:127–135. - PubMed
-
- Bazzoni F, Beutler B. The tumor necrosis factor ligand and receptor families. N Engl J Med. 1996;334:1717–1725. - PubMed
-
- Cardoni R L, Rottenberg M E, Segura E L. Increased production of reactive oxygen species by spleen cells from mice acutely infected with Trypanosoma cruzi. Cell Immunol. 1990;128:11–21. - PubMed
-
- Chandrasekar B, Melby P, Troyer D, Freeman G. Induction of proinflammatory cytokine expression in experimental acute chagasic cardiomyopathy. Biochem Biophys Res Commun. 1996;223:365–371. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
