

SMART, a simple modular architecture research tool: identification of signaling domains
- PMID: 9600884
- PMCID: PMC34487
- DOI: 10.1073/pnas.95.11.5857
SMART, a simple modular architecture research tool: identification of signaling domains
Abstract
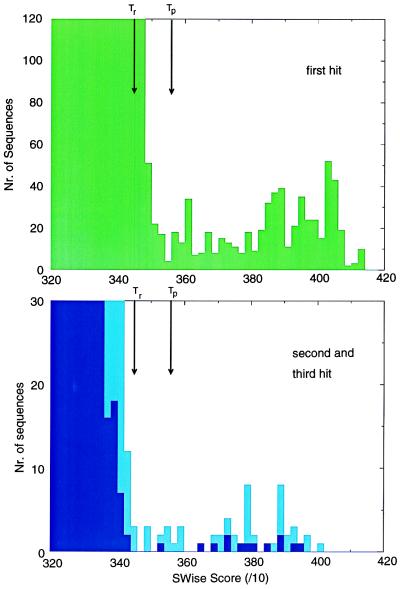
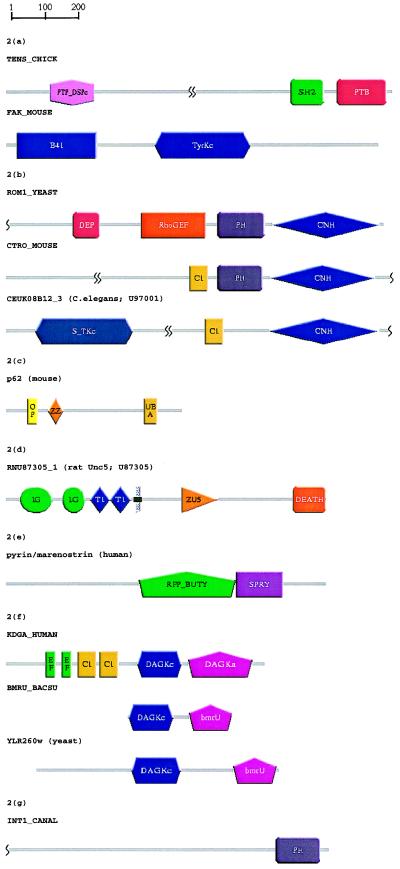
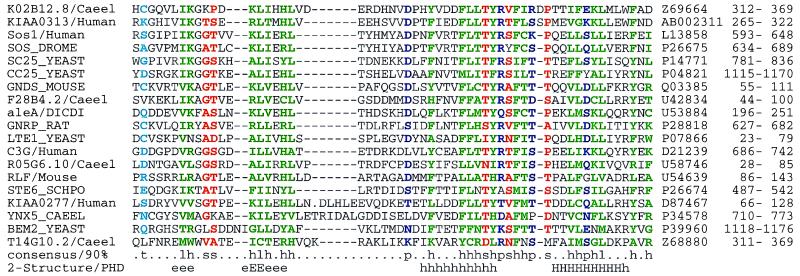
Accurate multiple alignments of 86 domains that occur in signaling proteins have been constructed and used to provide a Web-based tool (SMART: simple modular architecture research tool) that allows rapid identification and annotation of signaling domain sequences. The majority of signaling proteins are multidomain in character with a considerable variety of domain combinations known. Comparison with established databases showed that 25% of our domain set could not be deduced from SwissProt and 41% could not be annotated by Pfam. SMART is able to determine the modular architectures of single sequences or genomes; application to the entire yeast genome revealed that at least 6.7% of its genes contain one or more signaling domains, approximately 350 greater than previously annotated. The process of constructing SMART predicted (i) novel domain homologues in unexpected locations such as band 4.1-homologous domains in focal adhesion kinases; (ii) previously unknown domain families, including a citron-homology domain; (iii) putative functions of domain families after identification of additional family members, for example, a ubiquitin-binding role for ubiquitin-associated domains (UBA); (iv) cellular roles for proteins, such predicted DEATH domains in netrin receptors further implicating these molecules in axonal guidance; (v) signaling domains in known disease genes such as SPRY domains in both marenostrin/pyrin and Midline 1; (vi) domains in unexpected phylogenetic contexts such as diacylglycerol kinase homologues in yeast and bacteria; and (vii) likely protein misclassifications exemplified by a predicted pleckstrin homology domain in a Candida albicans protein, previously described as an integrin.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
