

Chaperone-facilitated copper binding is a property common to several classes of familial amyotrophic lateral sclerosis-linked superoxide dismutase mutants
- PMID: 9600970
- PMCID: PMC27707
- DOI: 10.1073/pnas.95.11.6361
Chaperone-facilitated copper binding is a property common to several classes of familial amyotrophic lateral sclerosis-linked superoxide dismutase mutants
Abstract
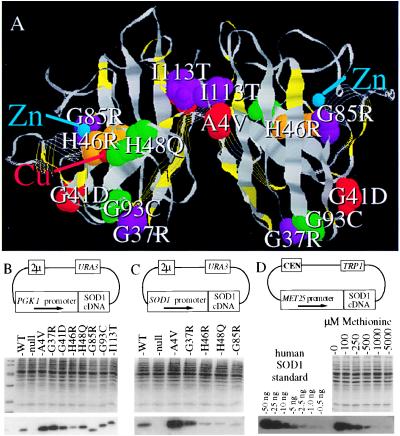
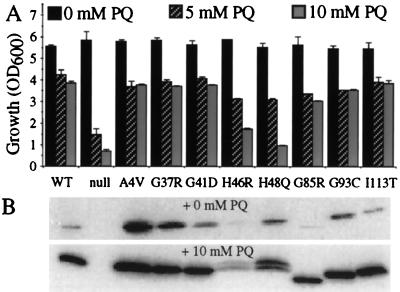
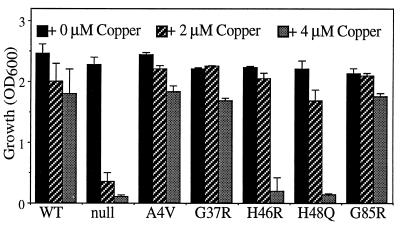
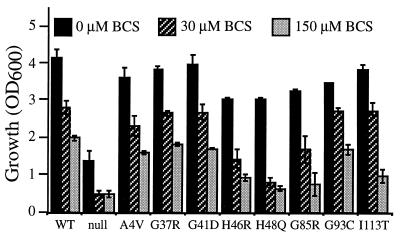
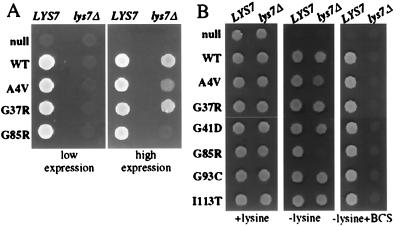
Mutations in Cu, Zn superoxide dismutase (SOD1) cause the neurodegenerative disease familial amyotrophic lateral sclerosis from an as-yet-unidentified toxic property(ies). Analysis in Saccharomyces cerevisiae of a broad range of human familial amyotrophic lateral sclerosis-linked SOD1 mutants (A4V, G37R, G41D, H46R, H48Q, G85R, G93C, and I113T) reveals one property common to these mutants (including two at residues that coordinate the catalytic copper): Each does indeed bind copper and scavenge oxygen-free radicals in vivo. Neither decreased copper binding nor decreased superoxide scavenging activity is a property shared by all mutants. The demonstration that shows that all mutants tested do bind copper under physiologic conditions supports a mechanism of SOD1 mutant-mediated disease arising from aberrant copper-mediated chemistry catalyzed by less tightly folded (and hence less constrained) mutant enzymes. The mutant enzymes also are shown to acquire the catalytic copper in vivo through the action of CCS, a specific copper chaperone for SOD1, which in turn suggests that a search for inhibitors of this SOD1 copper chaperone may represent a therapeutic avenue.
Figures
References
-
- Rosen D R, Siddique T, Patterson D, Figlewicz D A, Sapp P, Hentati A, Donaldson D, Goto J, O’Regan J P, Deng H X, et al. Nature (London) 1993;362:59–62. - PubMed
-
- Deng H-X, Hentati A, Tainer J A, Iqbal Z, Cayabyab A, Hung W-Y, Getzoff E D, Hu P, Herzfeldt B, Roos R P, et al. Science. 1993;261:1047–1051. - PubMed
-
- Wong P C, Pardo C A, Borchelt D R, Lee M K, Copeland N G, Jenkins N A, Sisodia S S, Cleveland D W, Price D L. Neuron. 1995;14:1105–1116. - PubMed
-
- Bruijn L I, Becher M W, Lee M K, Anderson K L, Jenkins N A, Copeland N G, Sisodia S S, Rothstein J D, Borchelt D R, Price D L, et al. Neuron. 1997;18:327–338. - PubMed
-
- Reaume A B, Elliott J L, Hoffman E K, Kowall N W, Ferrante R J, Siwek D F, Wilcox H M, Flood D G, Beal M F, Brown R H, et al. Nat Genet. 1996;13:43–47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
