

Precisely full length, circularizable, complementary RNA: an infectious form of potato spindle tuber viroid
- PMID: 9601006
- PMCID: PMC27879
- DOI: 10.1073/pnas.95.11.6560
Precisely full length, circularizable, complementary RNA: an infectious form of potato spindle tuber viroid
Abstract
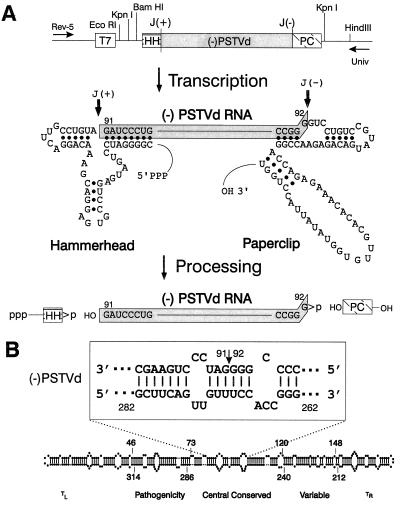
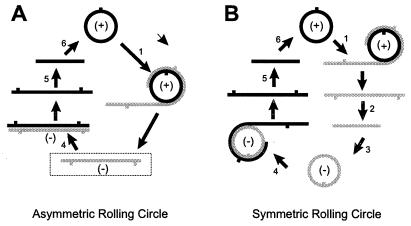
The replication of many viral and subviral pathogens as well as the amplification of certain cellular genes proceeds via a rolling circle mechanism. For potato spindle tuber (PSTVd) and related viroids, the possible role of a circular (-)strand RNA as a template for synthesis of (+)strand progeny is unclear. Infected plants appear to contain only multimeric linear (-)strand RNAs, and attempts to initiate infection with multimeric (-)PSTVd RNAs generally have failed. To examine critically the infectivity of monomeric (-)strand viroid RNAs, we have developed a ribozyme-based expression system for the production of precisely full length (-)strand RNAs whose termini are capable of undergoing facile circularization in vitro. Mechanical inoculation of tomato seedlings with electrophoretically purified (-)PSTVd RNA led to a small fraction of plants becoming infected whereas parallel assays with an analogous tomato planta macho viroid (-)RNA resulted in a much larger fraction of infected plants. Ribozyme-mediated production of (-)PSTVd RNA in transgenic plants led to the appearance of monomeric circular (-)PSTVd RNA and large amounts of (+)PSTVd progeny. No monomeric circular (-)PSTVd RNA could be detected in naturally infected plants by using either ribonuclease protection or electrophoresis under partially denaturing conditions. Although not a component of the normal replicative pathway, precisely full length (-)PSTVd RNA appears to contain all of the structural and regulatory elements necessary for initiation of viroid replication.
Figures
References
-
- Diener T O, editor. The Viroids. New York: Plenum; 1987.
-
- Semancik J S, editor. Viroids and Viroid-Like Pathogens. Boca Raton, FL: CRC; 1987.
-
- Branch A D, Robertson H D. Science. 1984;223:450–455. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
