

Towards a biocatalyst for (S)-styrene oxide production: characterization of the styrene degradation pathway of Pseudomonas sp. strain VLB120
- PMID: 9603811
- PMCID: PMC106275
- DOI: 10.1128/AEM.64.6.2032-2043.1998
Towards a biocatalyst for (S)-styrene oxide production: characterization of the styrene degradation pathway of Pseudomonas sp. strain VLB120
Erratum in
- Appl Environ Microbiol 1998 Sep;64(9):3546
Abstract
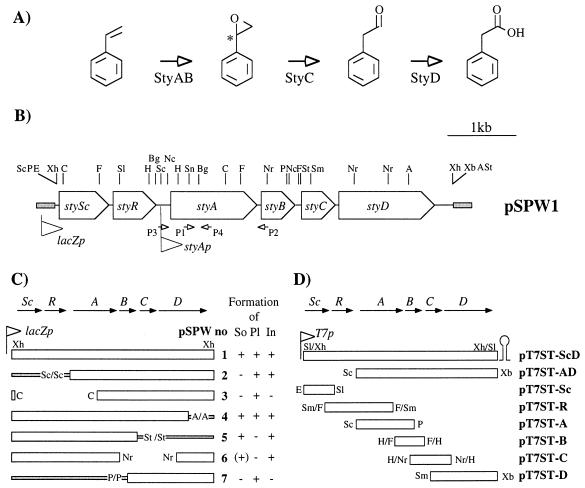
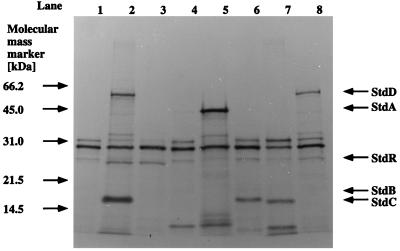
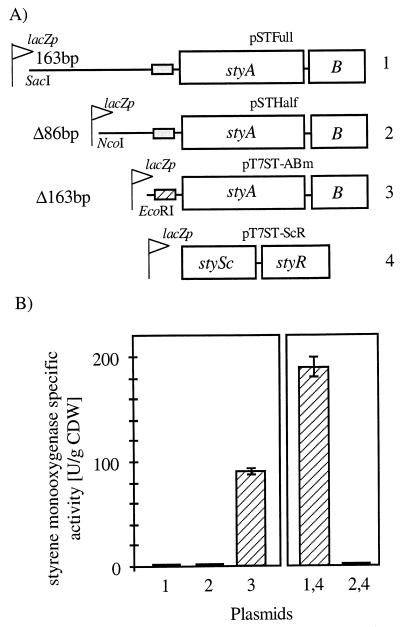
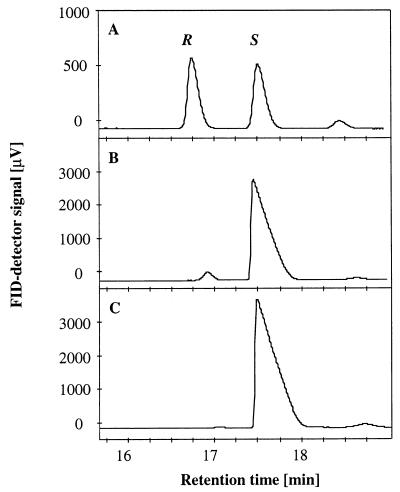
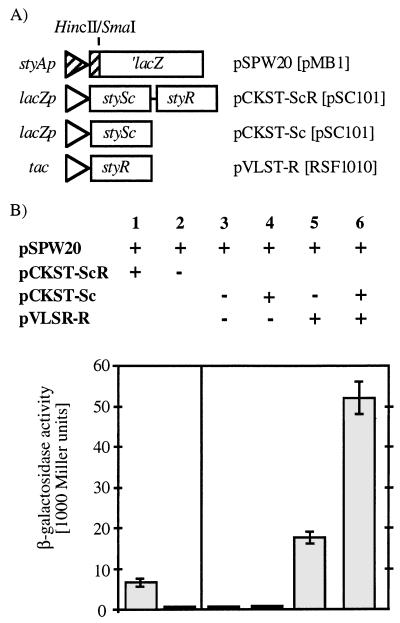
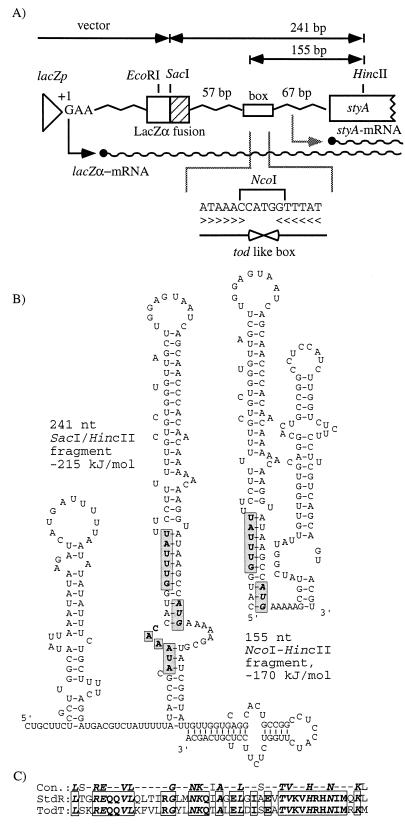
In order to design a biocatalyst for the production of optically pure styrene oxide, an important building block in organic synthesis, the metabolic pathway and molecular biology of styrene degradation in Pseudomonas sp. strain VLB120 was investigated. A 5.7-kb XhoI fragment, which contained on the same strand of DNA six genes involved in styrene degradation, was isolated from a gene library of this organism in Escherichia coli by screening for indigo formation. T7 RNA polymerase expression experiments indicated that this fragment coded for at least five complete polypeptides, StyRABCD, corresponding to five of the six genes. The first two genes encoded the potential carboxy-terminal part of a sensor, named StySc, and the complete response regulator StyR. Fusion of the putative styAp promoter to a lacZ reporter indicated that StySc and StyR together regulate expression of the structural genes at the transcriptional level. Expression of styScR also alleviated a block that prevented translation of styA mRNA when a heterologous promoter was used. The structural genes styA and styB produced a styrene monooxygenase that converted styrene to styrene oxide, which was then converted to phenylacetaldehyde by StyC. Sequence homology analysis of StyD indicated a probable function as a phenylacetaldehyde dehydrogenase. To assess the usefulness of the enzymes for the production of enantiomerically pure styrene oxide, we investigated the enantiospecificities of the reactions involved. Kinetic resolution of racemic styrene oxide by styrene oxide isomerase was studied with E. coli recombinants carrying styC, which converted styrene oxide at a very high rate but with only a slight preference for the S enantiomer. However, recombinants producing styrene monooxygenase catalyzed the formation of (S)-styrene oxide from inexpensive styrene with an excellent enantiomeric excess of more than 99% at rates up to 180 U g (dry weight) of cells-1.
Figures
References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Anthamatten D, Hennecke H. The regulatory status of the fixL- and fixJ-like genes in Bradyrhizobium japonicum may be different from that in Rhizobium meliloti. Mol Gen Genet. 1991;225:38–48. - PubMed
-
- Bond J A. Review of the toxicology of styrene. Crit Rev Toxicol. 1989;19:227–249. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
