

Characterization of the hca cluster encoding the dioxygenolytic pathway for initial catabolism of 3-phenylpropionic acid in Escherichia coli K-12
- PMID: 9603882
- PMCID: PMC107259
- DOI: 10.1128/JB.180.11.2915-2923.1998
Characterization of the hca cluster encoding the dioxygenolytic pathway for initial catabolism of 3-phenylpropionic acid in Escherichia coli K-12
Abstract
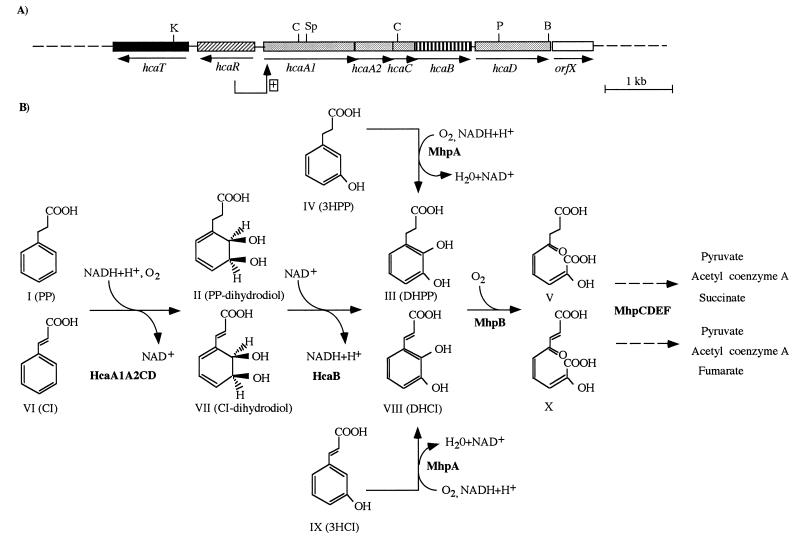
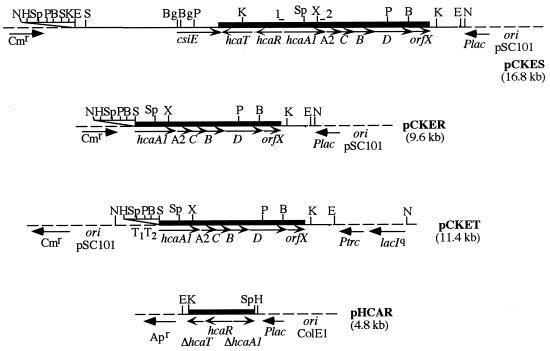
We have identified, cloned, and sequenced the hca cluster encoding the dioxygenolytic pathway for initial catabolism of 3-phenylpropionic acid (PP) in Escherichia coli K-12. This cluster maps at min 57.5 of the chromosome and is composed of five catabolic genes arranged as a putative operon (hcaA1A2CBD) and two additional genes transcribed in the opposite direction that encode a potential permease (hcaT) and a regulator (hcaR). Sequence comparisons revealed that while hcaA1A2CD genes encode the four subunits of the 3-phenylpropionate dioxygenase, the hcaB gene codes for the corresponding cis-dihydrodiol dehydrogenase. This type of catabolic module is homologous to those encoding class IIB dioxygenases and becomes the first example of such a catabolic cluster in E. coli. The inducible expression of the hca genes requires the presence of the hcaR gene product, which acts as a transcriptional activator and shows significant sequence similarity to members of the LysR family of regulators. Interestingly, the HcaA1A2CD and HcaB enzymes are able to oxidize not only PP to 3-(2,3-dihydroxyphenyl)propionate (DHPP) but also cinnamic acid (CI) to its corresponding 2, 3-dihydroxy derivative. Further catabolism of DHPP requires the mhp-encoded meta fission pathway for the mineralization of 3-hydroxyphenylpropionate (3HPP) (A. Ferrández, J. L. García, and E. Díaz, J. Bacteriol. 179:2573-2581, 1997). Expression in Salmonella typhimurium of the mhp genes alone or in combination with the hca cluster allowed the growth of the recombinant bacteria in 3-hydroxycinnamic acid (3HCI) and CI, respectively. Thus, the convergent mhp- and hca-encoded pathways are also functional in S. typhimurium, and they are responsible for the catabolism of different phenylpropanoid compounds (3HPP, 3HCI, PP, and CI) widely available in nature.
Figures

References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Asturias J A, Díaz E, Timmis K N. The evolutionary relationships of biphenyl dioxygenase from Gram-positive Rhodococcus globerulus P6 to multicomponent dioxygenases from Gram-negative bacteria. Gene. 1995;156:11–18. - PubMed
-
- Bachmann B J. Derivations and genotypes of some mutant derivatives of Escherichia coli K-12. In: Neidhardt F C, Ingraham J L, Low K B, Magasanik B, Schaechter M, Umbarger H E, editors. Escherichia coli and Salmonella thyphimurium: cellular and molecular biology. Washington, D.C: American Society for Microbiology; 1987. pp. 1190–1219.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
