

Phylogeny of dissimilatory sulfite reductases supports an early origin of sulfate respiration
- PMID: 9603890
- PMCID: PMC107267
- DOI: 10.1128/JB.180.11.2975-2982.1998
Phylogeny of dissimilatory sulfite reductases supports an early origin of sulfate respiration
Abstract
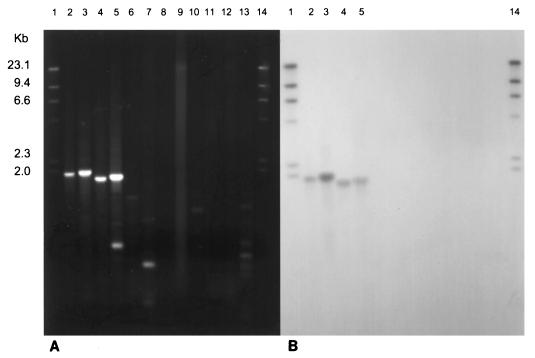
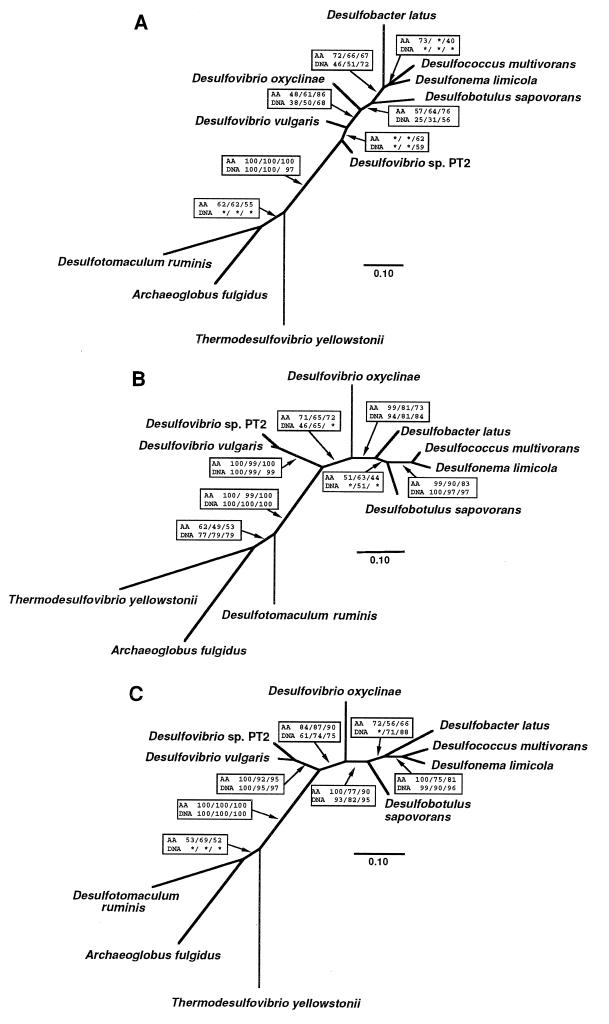
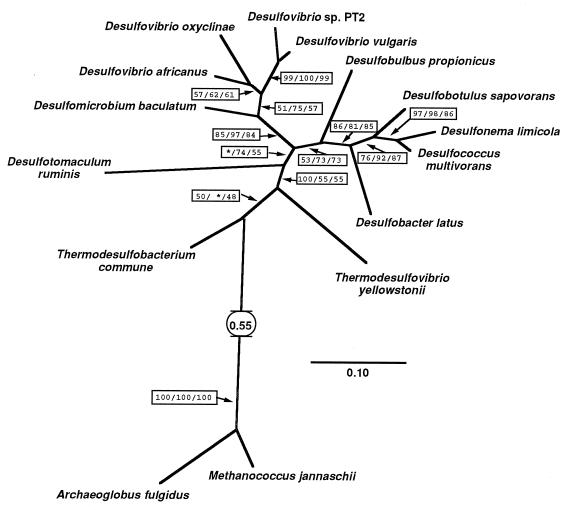
Microorganisms that use sulfate as a terminal electron acceptor for anaerobic respiration play a central role in the global sulfur cycle. Here, we report the results of comparative sequence analysis of dissimilatory sulfite reductase (DSR) genes from closely and distantly related sulfate-reducing organisms to infer the evolutionary history of DSR. A 1.9-kb DNA region encoding most of the alpha and beta subunits of DSR could be recovered only from organisms capable of dissimilatory sulfate reduction with a PCR primer set targeting highly conserved regions in these genes. All DNA sequences obtained were highly similar to one another (49 to 89% identity), and their inferred evolutionary relationships were nearly identical to those inferred on the basis of 16S rRNA. We conclude that the high similarity of bacterial and archaeal DSRs reflects their common origin from a conserved DSR. This ancestral DSR was either present before the split between the domains Bacteria, Archaea, and Eucarya or laterally transferred between Bacteria and Archaea soon after domain divergence. Thus, if the physiological role of the DSR was constant over time, then early ancestors of Bacteria and Archaea already possessed a key enzyme of sulfate and sulfite respiration.
Figures
References
-
- Achenbach-Richter L, Gupta R, Stetter K O, Woese C R. Were the original eubacteria thermophiles? Syst Appl Microbiol. 1987;9:34–39. - PubMed
-
- Adachi J, Hasegawa M. Computer science monographs, no. 28. MOLPHY version 2.3: programs for molecular phylogenetics based on maximum likelihood. Tokyo, Japan: Institute of Statistics and Mathematics; 1996.
-
- Arendsen A F, Verhagen M F J M, Wolbert R B G, Pierik A J, Stams A J M, Jetten M S M, Hagen W R. The dissimilatory sulfite reductase from Desulfosarcina variabilis is a desulforubidin containing uncoupled metalated sirohemes and S=9/2 iron-sulfur clusters. Biochemistry. 1993;32:10323–10330. - PubMed
-
- Boehringer Mannheim Corporation. Genius system user’s guide for membrane hybridization, version 3.0. Indianapolis, Ind: Boehringer Mannheim Corporation; 1994.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
