

Defining the interactions between intermediate filaments and desmosomes
- PMID: 9606214
- PMCID: PMC2137181
- DOI: 10.1083/jcb.141.5.1229
Defining the interactions between intermediate filaments and desmosomes
Abstract
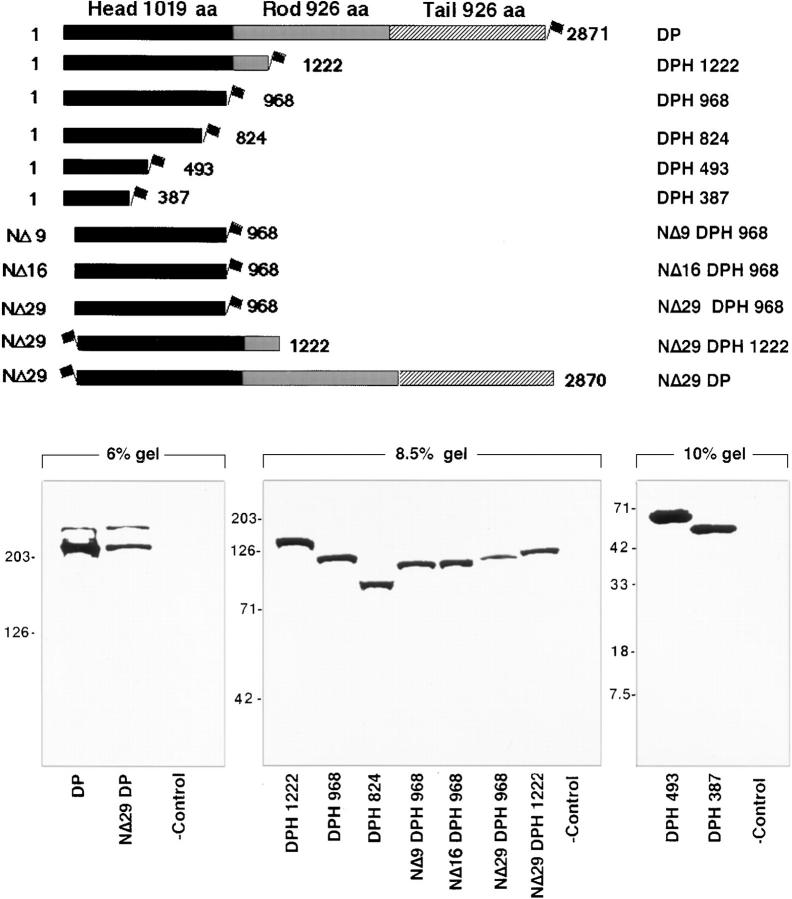
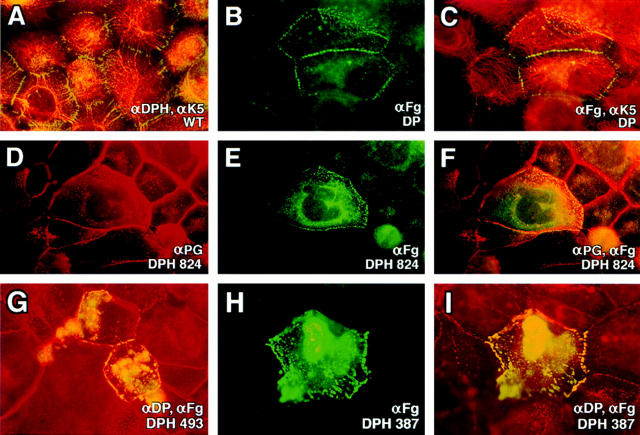
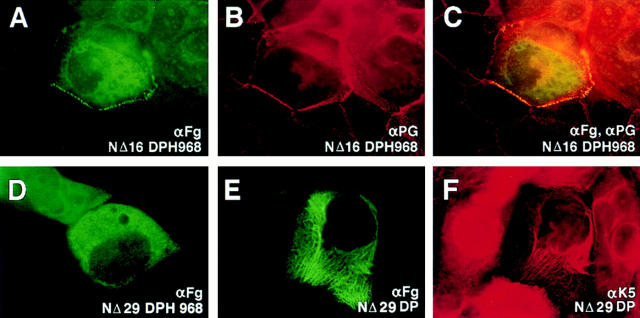
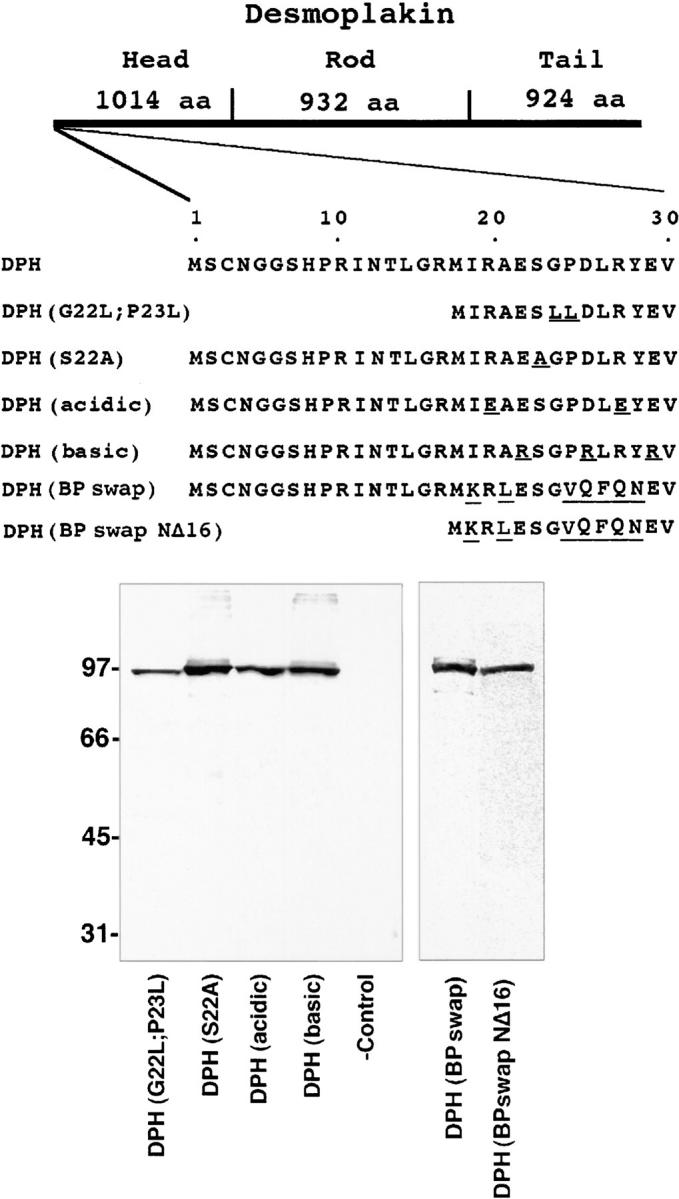
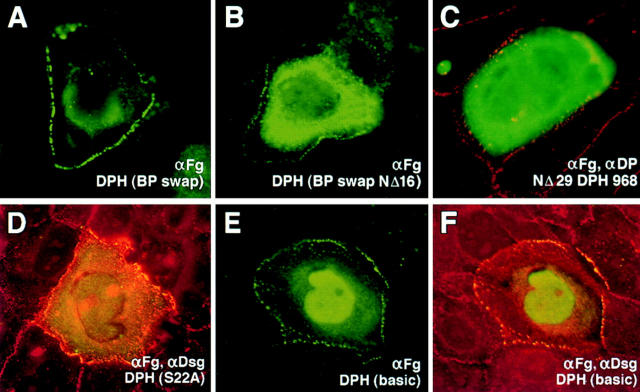
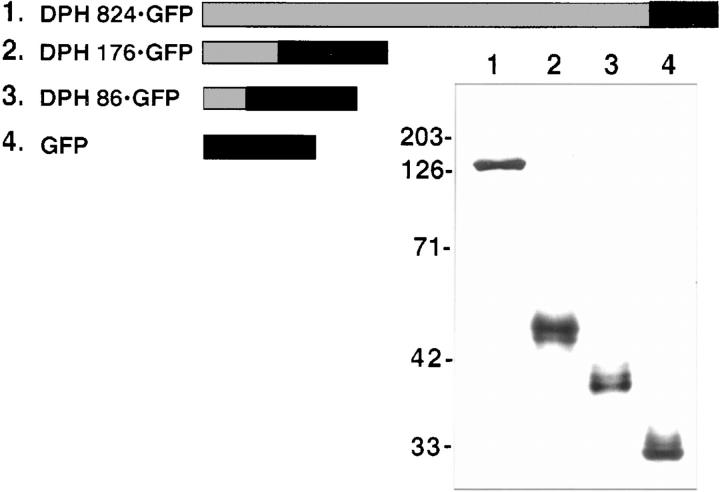
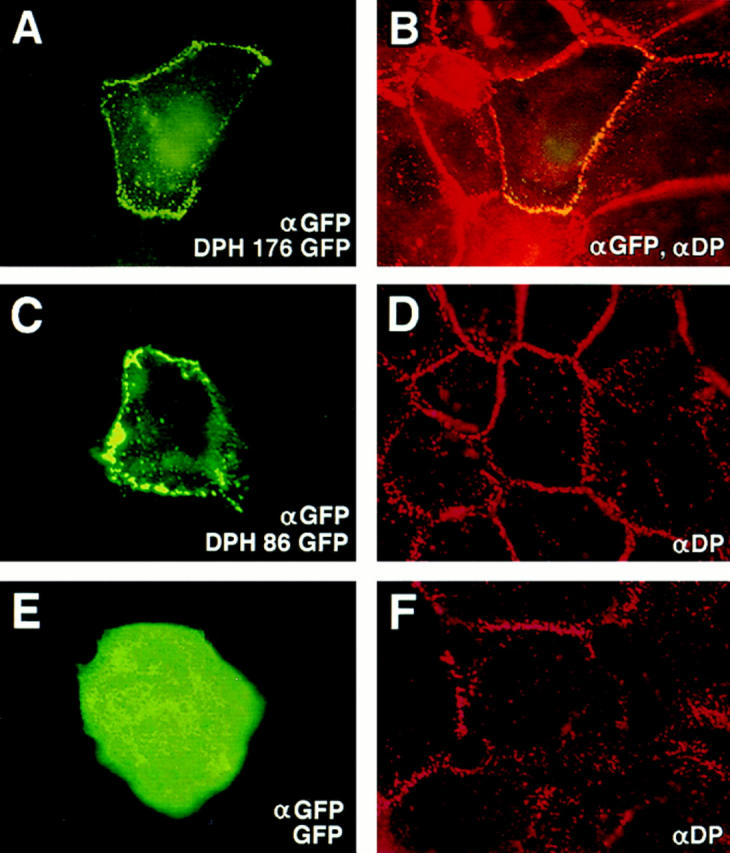
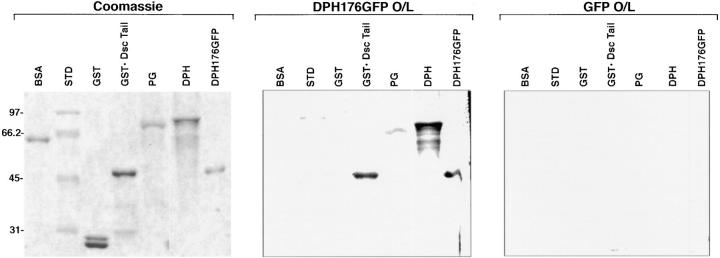
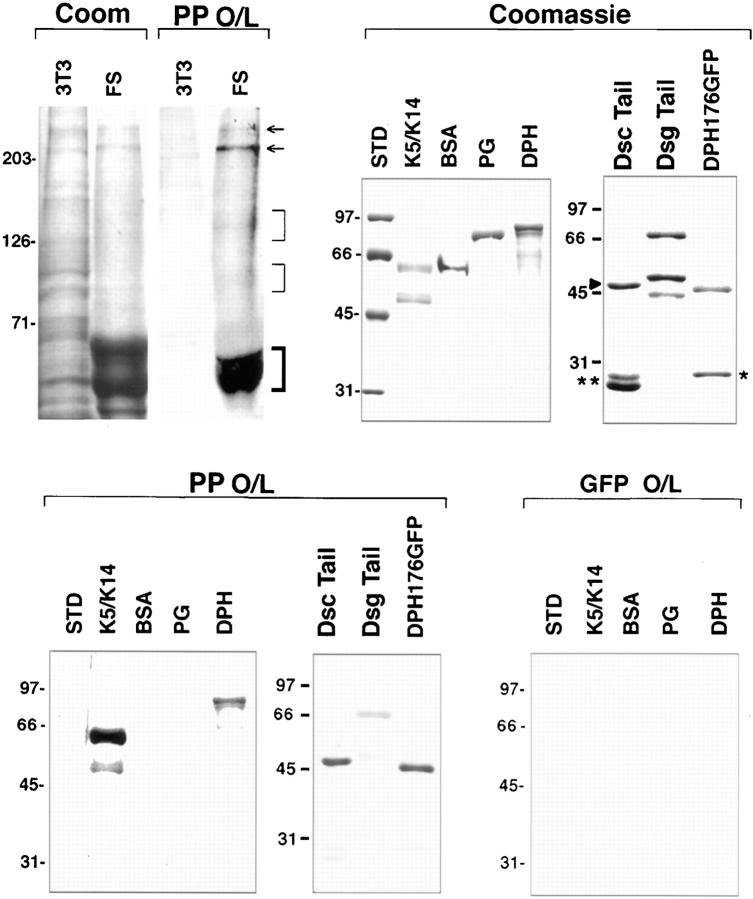
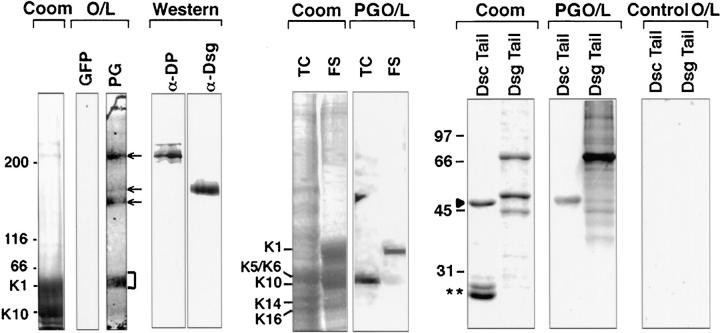
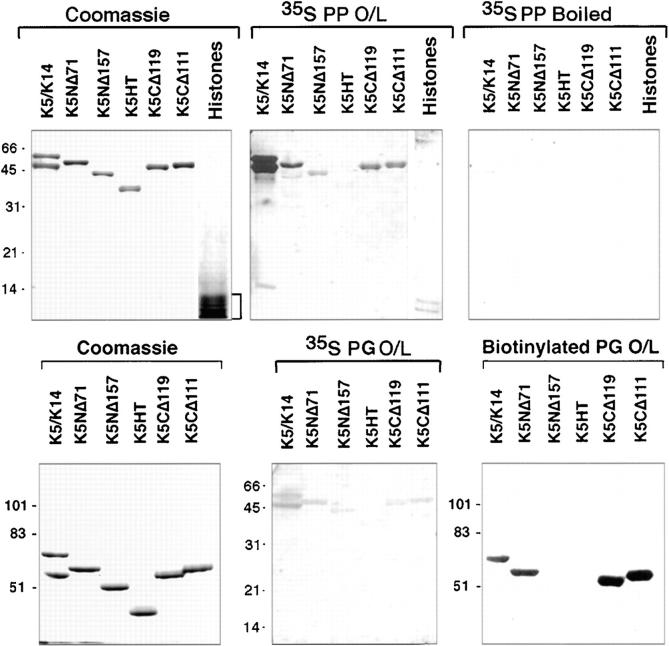
Desmoplakin (DP), plakoglobin (PG), and plakophilin 1 (PP1) are desmosomal components lacking a transmembrane domain, thus making them candidate linker proteins for connecting intermediate filaments and desmosomes. Using deletion and site-directed mutagenesis, we show that remarkably, removal of approximately 1% of DP's sequence obliterates its ability to associate with desmosomes. Conversely, when linked to a foreign protein, as few as 86 NH2-terminal DP residues are sufficient to target to desmosomes efficiently. In in vitro overlay assays, the DP head specifically associates with itself and with desmocollin 1a (Dsc1a). In similar overlay assays, PP1 binds to DP and Dsc1a, and to a lesser extent, desmoglein 1 (Dsg1), while PG binds to Dsg1 and more weakly to Dsc1a and DP. Interestingly, like DP, PG and PP1 associate with epidermal keratins, although PG is considerably weaker in its ability to do so. As judged by overlay assays, the amino terminal head domain of type II keratins appears to have a special importance in establishing these connections. Taken together, our findings provide new insights into the complexities of the links between desmosomes and intermediate filaments (IFs). Our results suggest a model whereby at desmosome sites within dividing epidermal cells, DP and PG anchor to desmosomal cadherins and to each other, forming an ordered array of nontransmembrane proteins that then bind to keratin IFs. As epidermal cells differentiate, PP1 is added as a molecular reinforcement to the plaque, enhancing anchorage to IFs and accounting at least partially for the increase in numbers and stability of desmosomes in suprabasal cells.
Figures
References
-
- Barth AI, Nathke IS, Nelson WJ. Cadherins, catenins and APC protein: interplay between cytoskeletal complexes and signaling pathways. Curr Opin Cell Biol. 1997;9:683–690. - PubMed
-
- Bierkamp C, McLaughlin KJ, Schwarz H, Huber O, Kemler R. Embryonic heart and skin defects in mice lacking plakoglobin. Dev Biol. 1996;180:780–785. - PubMed
-
- Bornslaeger EA, Corcoran CM, Stappenbeck TS, Green KJ. Breaking the connection: displacement of the desmosomal plaque protein desmoplakin from cell-cell interfaces disrupts anchorage of intermediate filament bundles and alters intercellular junction assembly. J Cell Biol. 1996;134:985–1001. - PMC - PubMed
-
- Candi E, Tarcsa E, Digiovanna JJ, Compton JG, Elias PM, Marekov LN, Steinert PM. A highly conserved lysine residue on the head domain of type II keratins is essential for the attachment of keratin intermediate filaments to the cornified cell envelope through isopeptide crosslinking by transglutaminases. Proc Natl Acad Sci USA. 1998;95:2067–2072. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
