

The UNI3 gene is required for assembly of basal bodies of Chlamydomonas and encodes delta-tubulin, a new member of the tubulin superfamily
- PMID: 9614175
- PMCID: PMC25351
- DOI: 10.1091/mbc.9.6.1293
The UNI3 gene is required for assembly of basal bodies of Chlamydomonas and encodes delta-tubulin, a new member of the tubulin superfamily
Abstract
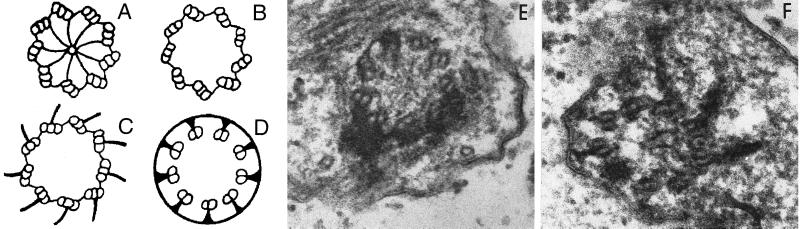
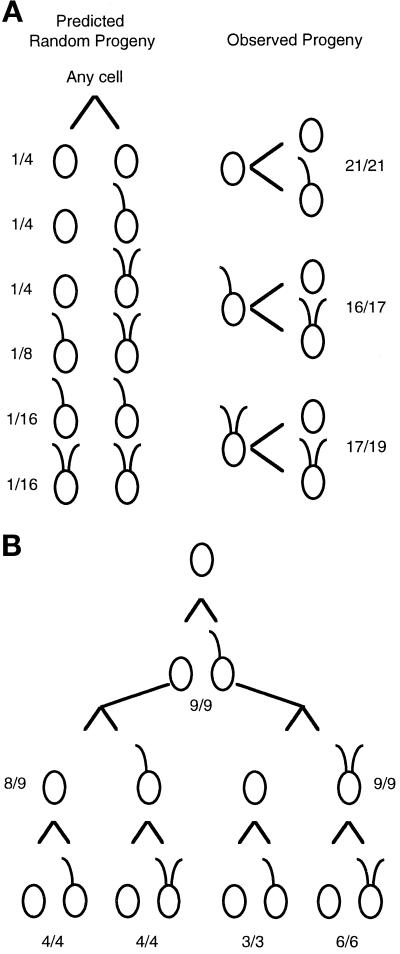
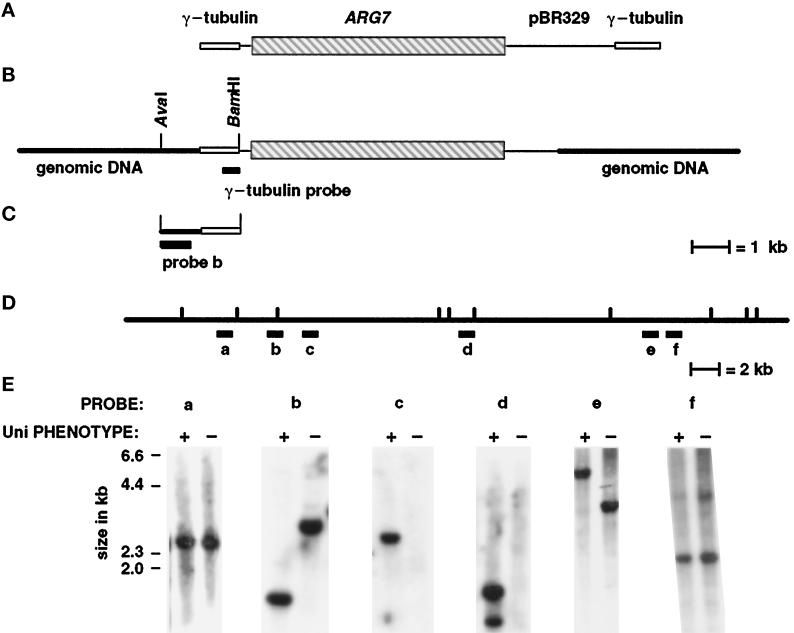
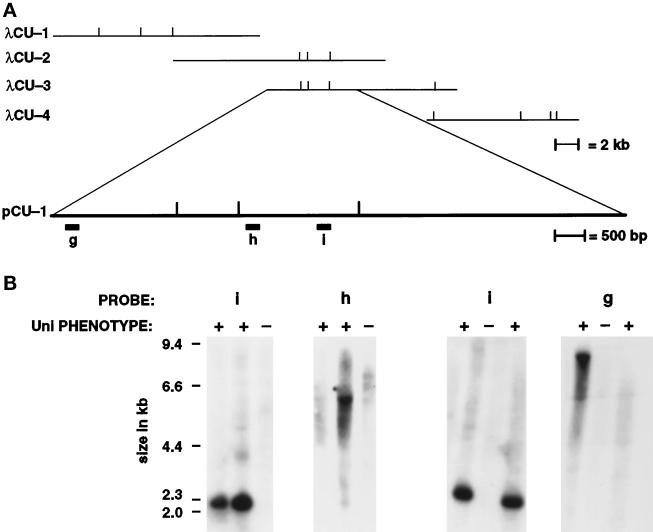
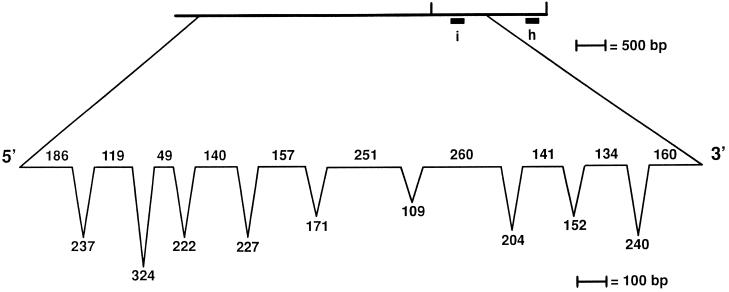
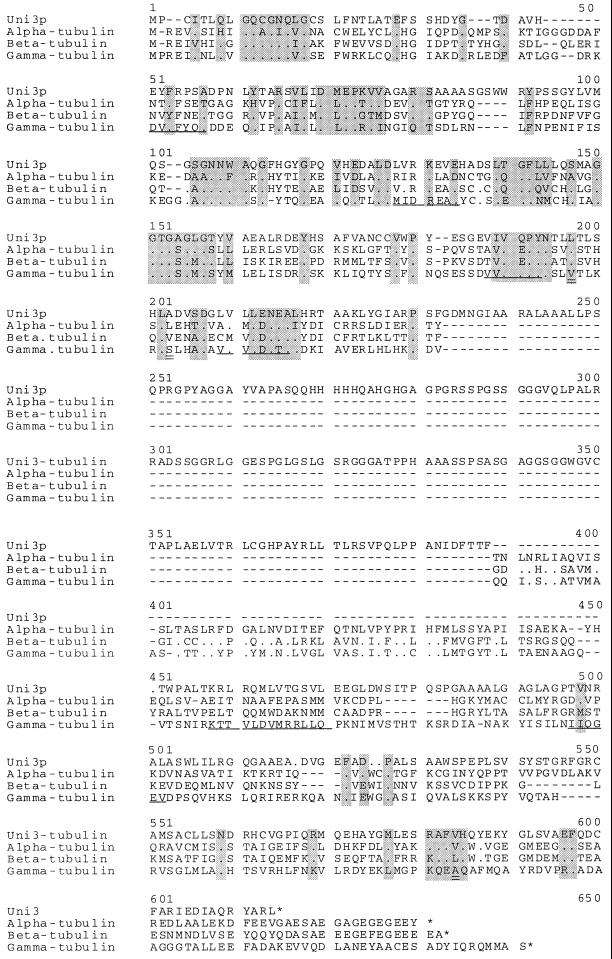
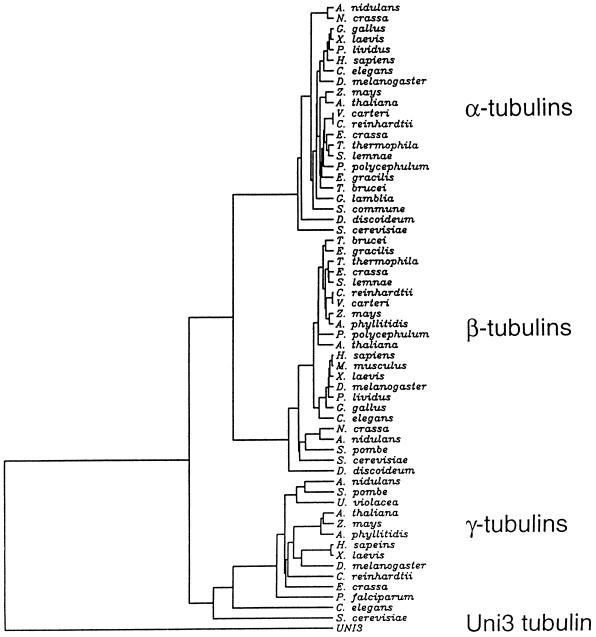
We have cloned the UNI3 gene in Chlamydomonas and find that it encodes a new member of the tubulin superfamily. Although Uni3p shares significant sequence identity with alpha-, beta-, and gamma-tubulins, there is a region of Uni3p that has no similarity to tubulins or other known proteins. Mutant uni3-1 cells assemble zero, one, or two flagella. Pedigree analysis suggests that flagellar number in uni3-1 cells is a function of the age of the cell. The uniflagellate uni3-1 cells show a positional phenotype; the basal body opposite the eyespot templates the single flagellum. A percentage of uni3-1 cells also fail to orient the cleavage furrow properly, and basal bodies have been implicated in the placement of cleavage furrows in Chlamydomonas. Finally when uni3-1 cells are observed by electron microscopy, doublet rather than triplet microtubules are observed at the proximal end of the basal bodies. We propose that the Uni3 tubulin is involved in both the function and cell cycle-dependent maturation of basal bodies/centrioles.
Figures
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Beech PL, Wetherbee R, Pickett-Heaps JD. Transformation of the flagella and associated flagellar components during cell division in the Coccolithophorid Pleurochysis carterie. Protoplasma. 1988;145:37–46.
-
- Burns RG. Identification of two new members of the tubulin family. Cell Motil Cytoskeleton. 1995;31:255–258. - PubMed
-
- Cabral F, Schibler M, Kuriyama R, Abraham I, Whitfield C. Genetic analysis of microtubule function in CHO cells. In: Borisy GG, Cleveland DW, Murphy DB, editors. Molecular Biology of the Cytoskeleton. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1984. pp. 305–317.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
