

Identification of osteopontin as a novel ligand for the integrin alpha8 beta1 and potential roles for this integrin-ligand interaction in kidney morphogenesis
- PMID: 9614184
- PMCID: PMC25364
- DOI: 10.1091/mbc.9.6.1425
Identification of osteopontin as a novel ligand for the integrin alpha8 beta1 and potential roles for this integrin-ligand interaction in kidney morphogenesis
Abstract
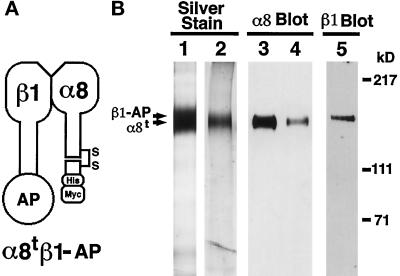
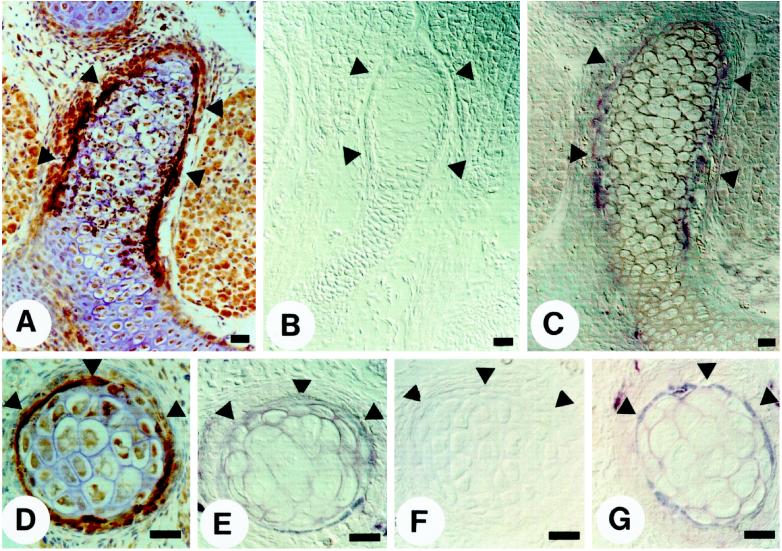
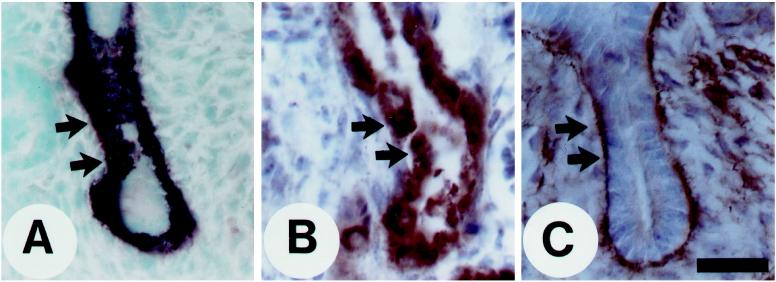
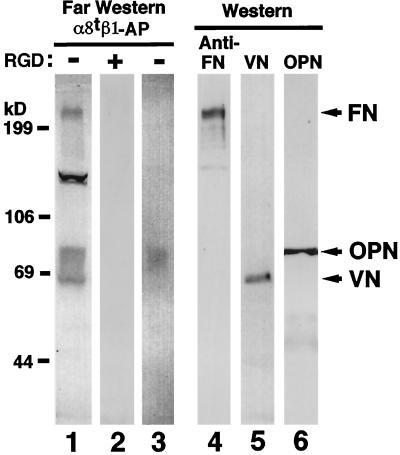
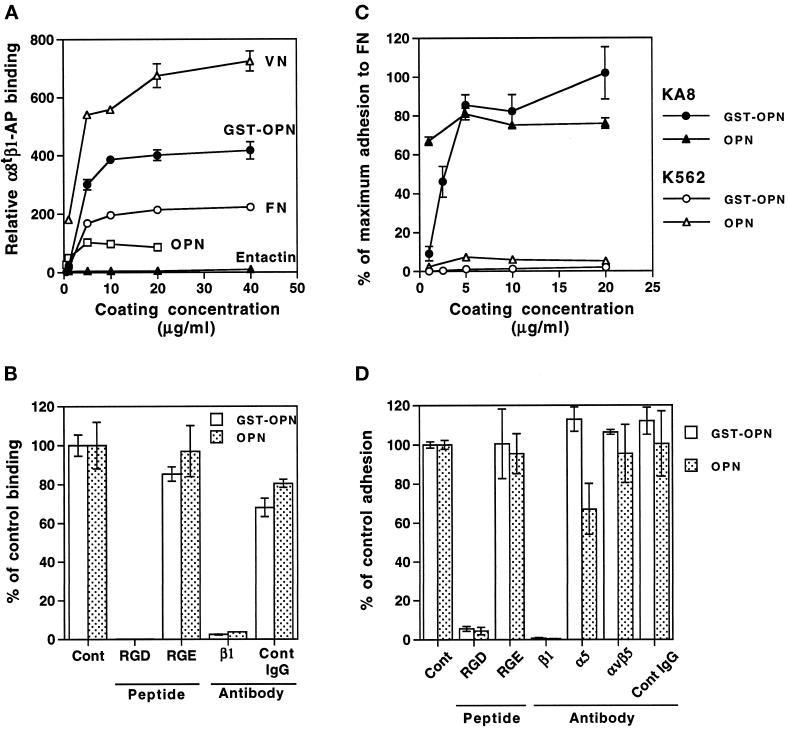
Epithelio-mesenchymal interactions during kidney organogenesis are disrupted in integrin alpha8 beta1-deficient mice. However, the known ligands for integrin alpha8 beta1-fibronectin, vitronectin, and tenascin-C-are not appropriately localized to mediate all alpha8 beta1 functions in the kidney. Using a method of general utility for determining the distribution of unknown integrin ligands in situ and biochemical characterization of these ligands, we identified osteopontin (OPN) as a ligand for alpha8 beta1. We have coexpressed the extracellular domains of the mouse alpha8 and beta1 integrin subunits as a soluble heterodimer with one subunit fused to alkaline phosphatase (AP) and have used the alpha8 beta1-AP chimera as a histochemical reagent on sections of mouse embryos. Ligand localization with alpha8 beta1-AP in developing bone and kidney was observed to be overlapping with the distribution of OPN. In "far Western" blots of mouse embryonic protein extracts, bands were detected with sizes corresponding to fibronectin, vitronectin, and unknown proteins, one of which was identical to the size of OPN. In a solid-phase binding assay we demonstrated that purified OPN binds specifically to alpha8 beta1-AP. Cell adhesion assays using K562 cells expressing alpha8 beta1 were used to confirm this result. Together with a recent report that anti-OPN antibodies disrupt kidney morphogenesis, our results suggest that interactions between OPN and integrin alpha8 beta1 may help regulate kidney development and other morphogenetic processes.
Figures
References
-
- Chen J, Zhang Q, McCulloch CAG, Sodek J. Immunohistochemical localization of bone sialoprotein in foetal procine bone tissues: comparisons with secreted phosphoprotein 1 (SPP-1, osteopontin) and SPARC (osteonectin) Histochem J. 1991;23:281–289. - PubMed
-
- Cheng H-J, Flanagan JG. Identification and cloning of ELF-1, a developmentally expressed ligand for the Mek4 and Sek receptor tyrosine kinases. Cell. 1994;79:157–168. - PubMed
-
- Clover J, Dodds RA, Gowen M. Integrin subunit expression by human osteoblasts and osteoclasts in situ and in culture. J Cell Sci. 1992;103:267–271. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
