

Regulation of X-chromosome inactivation in development in mice and humans
- PMID: 9618446
- PMCID: PMC98919
- DOI: 10.1128/MMBR.62.2.362-378.1998
Regulation of X-chromosome inactivation in development in mice and humans
Abstract
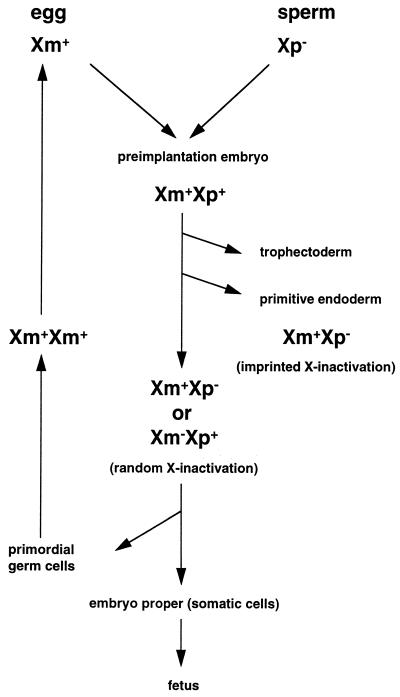
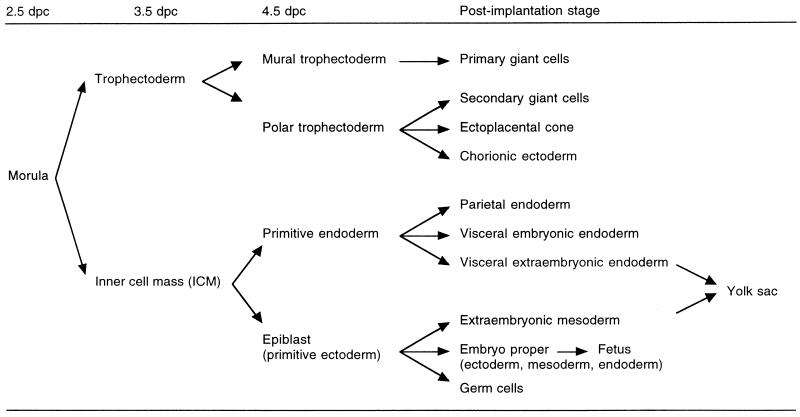
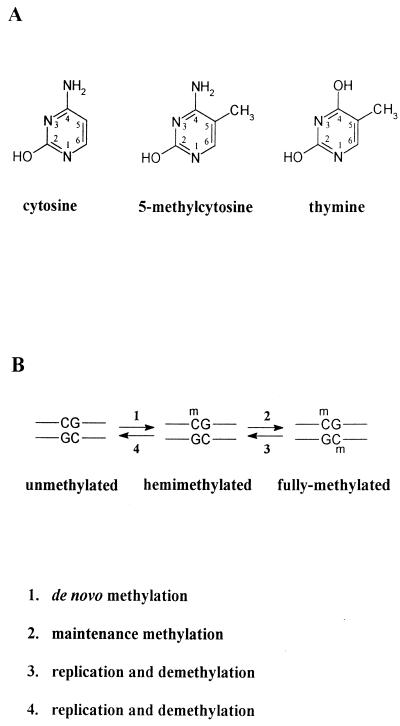
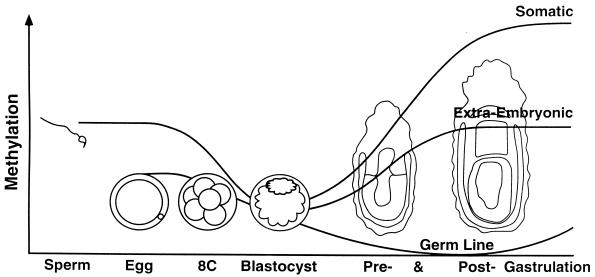
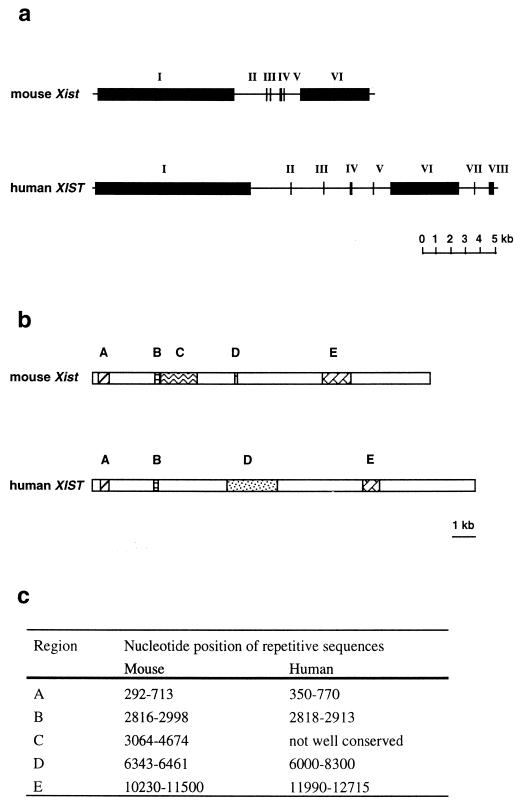
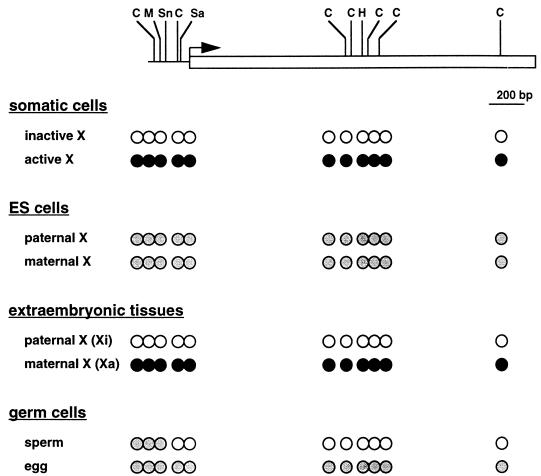
Dosage compensation for X-linked genes in mammals is accomplished by inactivating one of the two X chromosomes in females. X-chromosome inactivation (XCI) occurs during development, coupled with cell differentiation. In somatic cells, XCI is random, whereas in extraembryonic tissues, XCI is imprinted in that the paternally inherited X chromosome is preferentially inactivated. Inactivation is initiated from an X-linked locus, the X-inactivation center (Xic), and inactivity spreads along the chromosome toward both ends. XCI is established by complex mechanisms, including DNA methylation, heterochromatinization, and late replication. Once established, inactivity is stably maintained in subsequent cell generations. The function of an X-linked regulatory gene, Xist, is critically involved in XCI. The Xist gene maps to the Xic, it is transcribed only from the inactive X chromosome, and the Xist RNA associates with the inactive X chromosome in the nucleus. Investigations with Xist-containing transgenes and with deletions of the Xist gene have shown that the Xist gene is required in cis for XCI. Regulation of XCI is therefore accomplished through regulation of Xist. Transcription of the Xist gene is itself regulated by DNA methylation. Hence, the differential methylation of the Xist gene observed in sperm and eggs and its recognition by protein binding constitute the most likely mechanism regulating imprinted preferential expression of the paternal allele in preimplantation embryos and imprinted paternal XCI in extraembryonic tissues. This article reviews the mechanisms underlying XCI and recent advances elucidating the functions of the Xist gene in mice and humans.
Figures
References
-
- Adams R L P, Burdon R H, McKinnon K, Rinaldi A. Stimulation of de novo methylation following limited proteolysis of mouse ascites DNA methylase. FEBS Lett. 1983;163:194–198. - PubMed
-
- Adams R L P. Eukaryotic DNA methyltransferases—structure and function. Bioessays. 1995;17:139–145. - PubMed
-
- Antequera F, Boyes J, Bird A P. High levels of de novo methylation and altered chromatin structure at CpG islands in cell lines. Cell. 1990;62:503–514. - PubMed
-
- Ariel M, Robinson E, McCarrey J R, Cedar H. Gamete-specific methylation correlates with imprinting of the murine Xist gene. Nat Genet. 1995;9:312–315. - PubMed
-
- Barlow D P, Stoger R, Herrmann B G, Saito K, Schweiter N. The mouse insulin-like growth factor type-2 receptor is imprinted and closely linked to the Tme locus. Nature. 1991;349:84–87. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
