

Multistep signaling requirements for pituitary organogenesis in vivo
- PMID: 9620855
- PMCID: PMC316866
- DOI: 10.1101/gad.12.11.1691
Multistep signaling requirements for pituitary organogenesis in vivo
Abstract
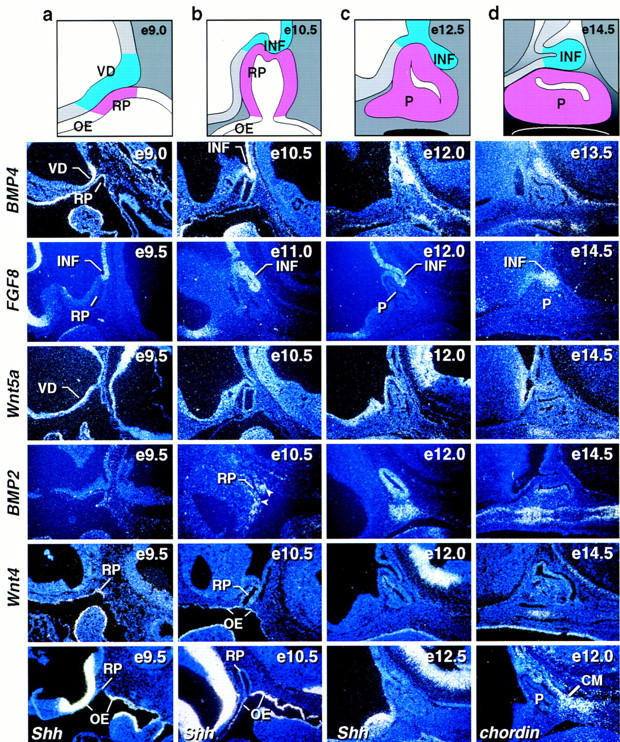
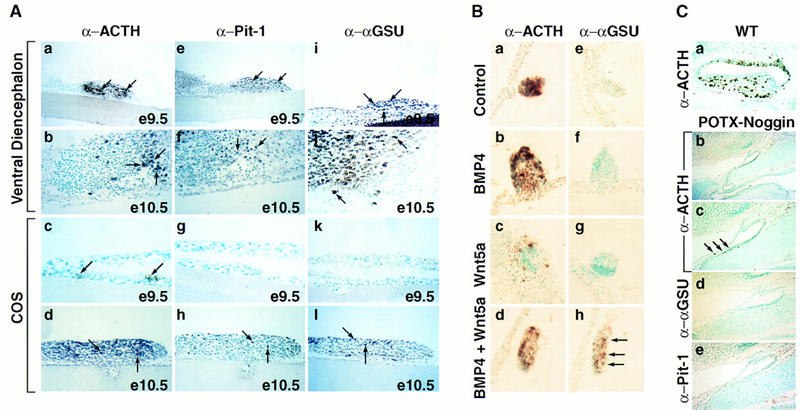
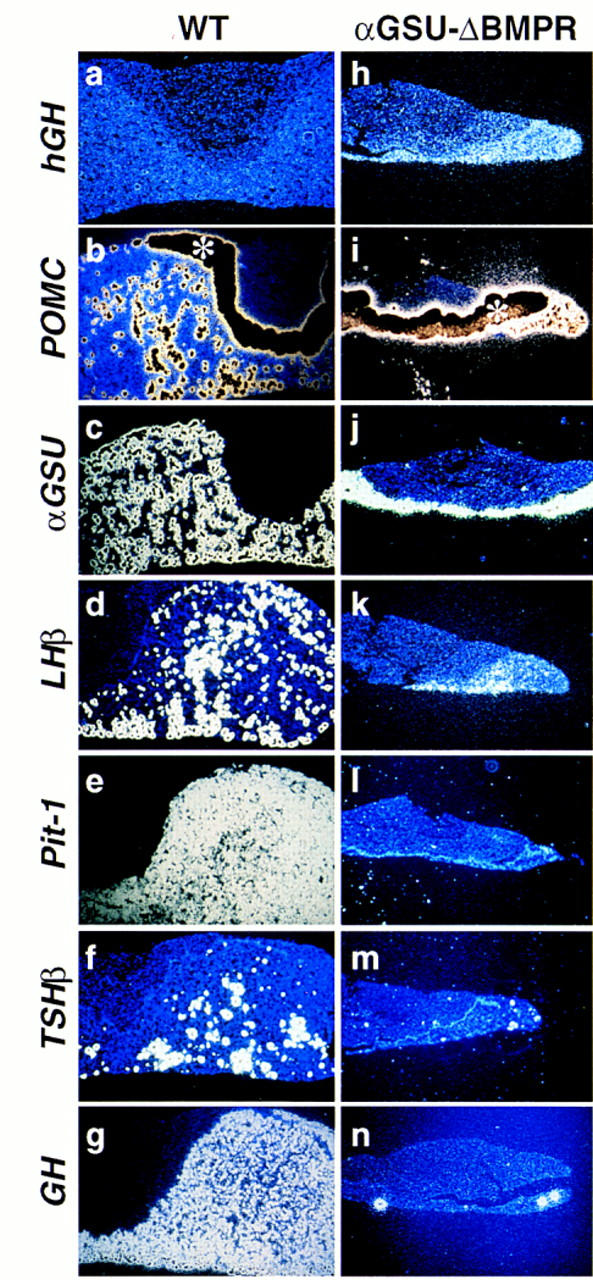
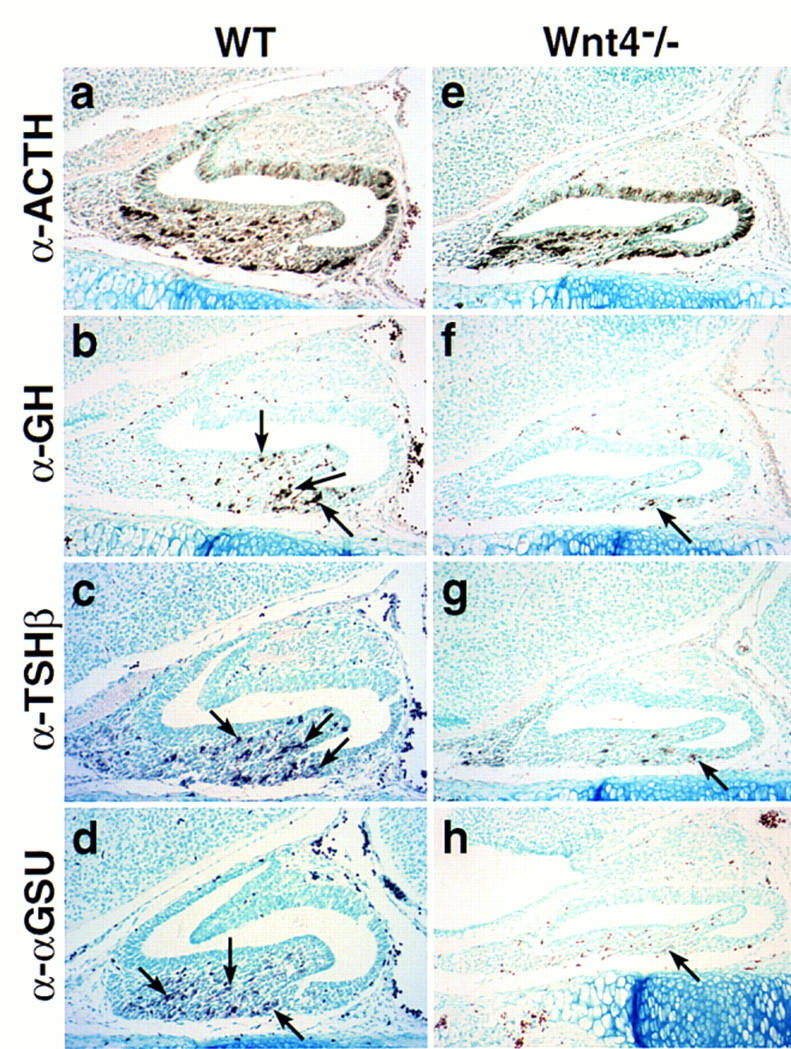
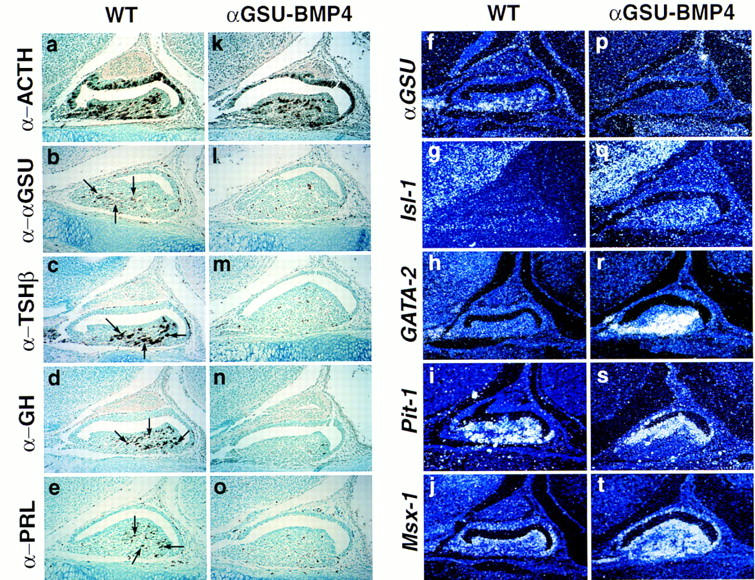
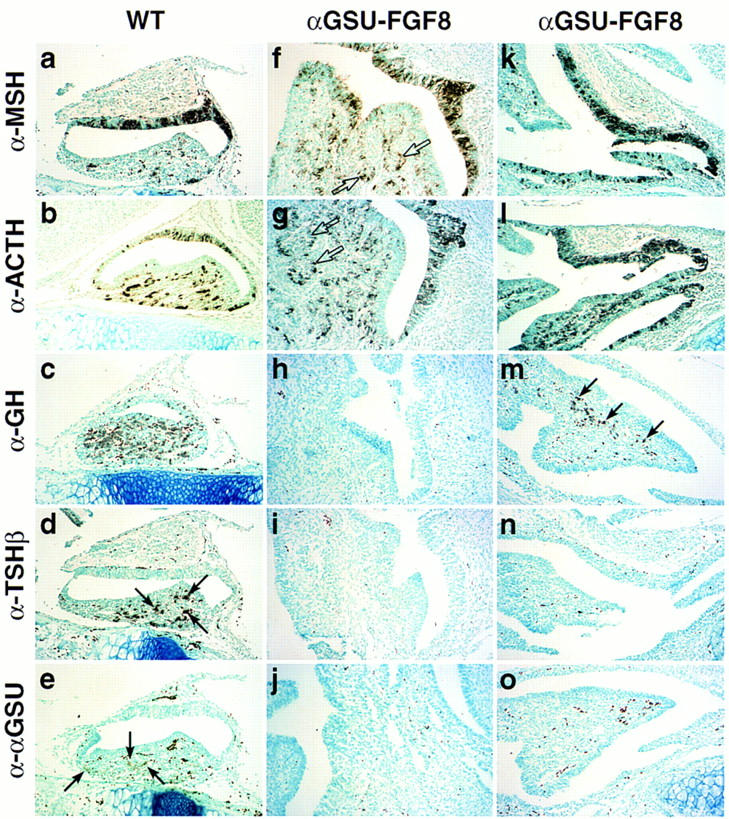
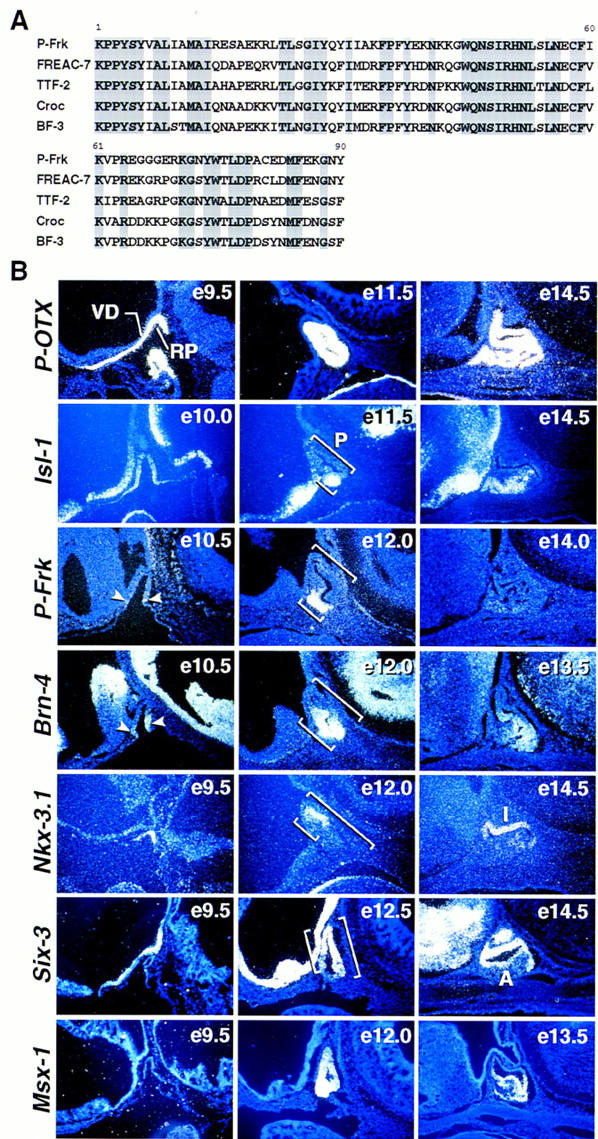
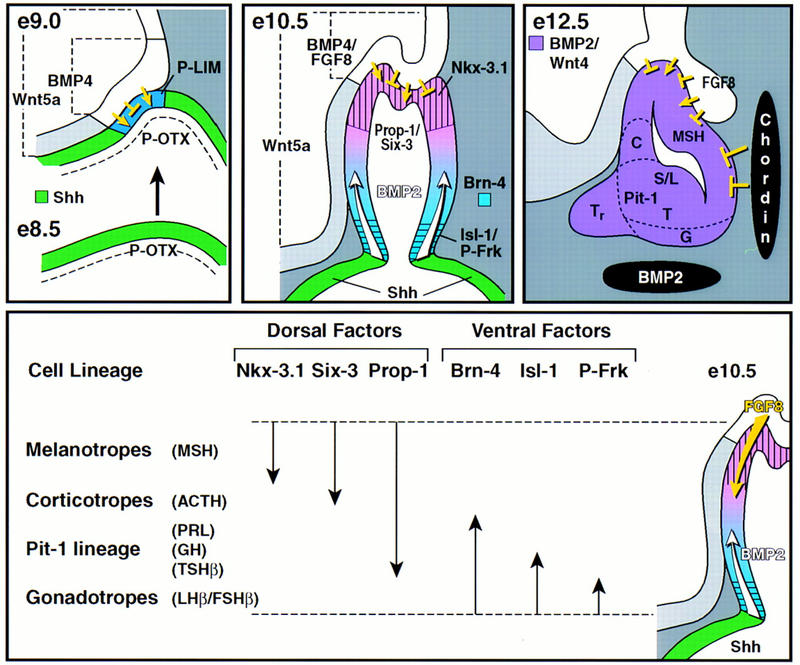
During development of the mammalian pituitary gland specific hormone-producing cell types, critical in maintaining homeostasis, emerge in a spatially and temporally specific fashion from an ectodermal primordium. We have investigated the molecular basis of generating diverse pituitary cell phenotypes from a common precursor, providing in vivo and in vitro evidence that their development involves three sequential phases of signaling events and the action of a gradient at an ectodermal boundary. In the first phase, the BMP4 signal from the ventral diencephalon, expressing BMP4, Wnt5a, and FGF8, represents a critical dorsal neuroepithelial signal for pituitary organ commitment in vivo. Subsequently, a BMP2 signal emanates from a ventral pituitary organizing center that forms at the boundary of a region of oral ectoderm in which Shh expression is selectively excluded. This BMP2 signal together with a dorsal FGF8 signal, appears to create opposing activity gradients that are suggested to generate overlapping patterns of specific transcription factors underlying cell lineage specification events, whereas Wnt4 is needed for the expansion of ventral pituitary cell phenotypes. In the third phase, temporally specific loss of the BMP2 signal is required to allow terminal differentiation. The consequence of these sequential organ and cellular determination events is that each of the hormone-producing pituitary cell types-gonadotropes, thyrotropes, somatotropes, lactotropes, corticotropes, and melanotropes-appear to be determined, in a ventral-to-dorsal gradient, respectively.
Figures
References
-
- Basler K, Struhl G. Compartment boundaries and the control of Drosophila limb pattern by hedgehog protein. Nature. 1994;368:208–214. - PubMed
-
- Bellusci S, Henderson R, Winnier G, Oikawa T, Hogan BL. Evidence from normal expression and targeted misexpression that bone morphogenetic protein (Bmp-4) plays a role in mouse embryonic lung morphogenesis. Development. 1996;122:1693–1702. - PubMed
-
- Bellusci S, Furuta Y, Rush MG, Henderson R, Winnier G, Hogan BL. Involvement of Sonic hedgehog (Shh) in mouse embryonic lung growth and morphogenesis. Development. 1997;124:53–63. - PubMed
-
- Bieberich C, Fujita K, He W, Jay G. Prostate-specific and androgen-dependent expression of a novel homeobox gene. J Biol Chem. 1996;271:31779–31782. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases