

Effects of carbon source on expression of F0 genes and on the stoichiometry of the c subunit in the F1F0 ATPase of Escherichia coli
- PMID: 9620972
- PMCID: PMC107823
- DOI: 10.1128/JB.180.12.3205-3208.1998
Effects of carbon source on expression of F0 genes and on the stoichiometry of the c subunit in the F1F0 ATPase of Escherichia coli
Abstract
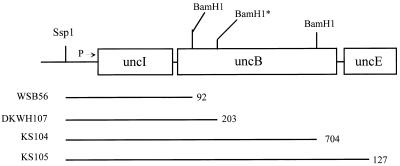
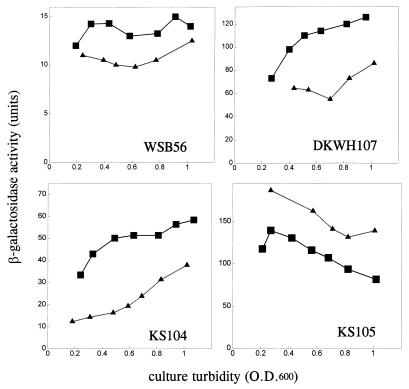
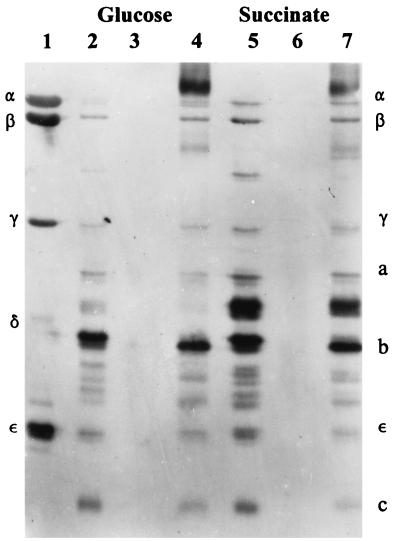
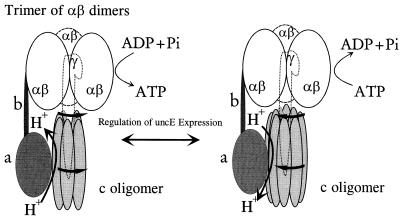
Expression of the genes for the membrane-bound F0 sector of the Escherichia coli F1F0 proton-translocating ATPase can respond to changes in metabolic conditions, and these changes are reflected in alterations in the subunit stoichiometry of the oligomeric F0 proton channel. Transcriptional and translational lacZ fusions to the promoter and to two F0 genes show that, during growth on the nonfermentable carbon source succinate, transcription of the operon and translation of uncB, encoding the a subunit of F0, are higher than during growth on glucose. In contrast, translation of the uncE gene, encoding the c subunit of F0, is higher during growth on glucose than during growth on succinate. Translation rates of both uncB and uncE change as culture density increases, but transcription rates do not. Quantitation of the c stoichiometry shows that more c subunits are assembled into the F1F0 ATPase in cells grown on glucose than in cells grown on succinate. E. coli therefore appears to have a mechanism for regulating the composition and, presumably, the function of the ATPase in response to metabolic circumstances.
Figures
References
-
- Arai H, Terres G, Pink S, Forgac M J. Topography and subunit stoichiometry of the coated vesicle proton pump. J Biol Chem. 1988;263:8796–8802. - PubMed
-
- Cross R L, Taiz L. Gene duplication as a means for altering H+/ATP ratios during the evolution of FoF1 ATPases and synthases. FEBS Lett. 1990;259:227–229. - PubMed
-
- Foster D L, Fillingame R H. Energy-transducing H+-ATPase of Escherichia coli. J Biol Chem. 1979;254:8230–8236. - PubMed
-
- Foster D L, Fillingame R H. Stoichiometry of subunits in the H+-ATPase complex of Escherichia coli. J Biol Chem. 1982;257:2009–2015. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
